行政院國家科學委員會專題研究計畫 期中進度報告
食用菇類誘導人類白血病 U937 細胞分化、熟成作用、蛋白
質體學分析及其對鼠皮下轉植 CT26 腫瘤之抑制作用(1/3)
計畫類別: 個別型計畫 計畫編號: NSC92-2313-B-002-047- 執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日 執行單位: 國立臺灣大學食品科技研究所 計畫主持人: 張鴻民 報告類型: 精簡報告 處理方式: 本計畫可公開查詢 中 華 民 國 93 年 5 月 21 日行政院國家科學委員會補助專題研究計畫成果報告
2 食用菇類誘導人類白血病 U937 細胞分化、成熟作用、蛋白質體學分析及其 對鼠皮下轉植 CT26 腫瘤之抑制作用 (1/3) 4 計畫類別:個別型計畫 6 計畫編號: NSC93-2313-B-002-047 執行期限:92/08/01- 93/07/31 8 計畫主持人:張鴻民 計畫參與人員:陳暄諭,陳珮宣 10 執行單位:台灣大學食品科技研究所 12 中華民國九十三年五月二十一日 14中文摘要
含生理活性成分菇類之攝取在生物體內有助於單核免疫細胞之活化作 2
用而提升抗腫瘤效果。經由飽和硫胺沈澱及 Sephacryl S-300 膠體過濾後所得
之柳松菇(Agrocybe aegerita, AA)與鴻喜菇(Hypsizigus mamoreus, HM)
4 純化蛋白質(5-200 µg/mL)與人類周邊血液分離而來之免疫單核球經過 1 天培 養後製備條件培養液(MNC-CM)。以 5-50 µg/mL 之 AA 或 HM 蛋白質所製備 6 之 MNC-CM 來共同培養 U937 細胞時發現,可抑制 45-90 % U937 之生長,而 直接以相當濃度菇類蛋白質來對血癌細胞作用時,則抑制作用不顯著,因 8 而推測單核球細胞之活化為主要原因。此外,分析 cytoplasmic superoxide 時 發現 25 µg/mL AA 或 HM 蛋白質可分別有 53 and 66 % 之 NBT (nitroblue 10 tetrazolium) 反應。此外,以 5-25 µg/mL 菇類蛋白質製備之 MNC-CM 可表現 出 U937 細胞之增加誘導成熟效果,誘導成熟之 U937 細胞亦有吞噬作用, 12 其吞噬能力可達33 %。成熟 U937 細胞之表面抗原(CD)表現方面,以 50 µg/mL AA 或 HM 蛋白質所製備之 MNC-CM 可表現出 50-70 %之 CD11b, CD14 and CD68 14 表現率。50 µg/mL AA 蛋白質所製備之 MNC-CM 以 ELISA 檢測得知含有高濃
度之 interleukine (IL)-1β (1120 pg/mL)、tumor necrosis factor (TNF)-α (1235 pg/mL), 16
及 320 pg/mL Granulocyte-macrophage colony-stimulating factor (GM-CSF)。 18
Abstract
Activation of mononuclear cells is supposed to be relevant to the antitumor effects in 2
vivo and is possibly influenced by high intake of dietary mushrooms containing some bioactive proteins. Purified protein fractions from cold-water extracts of Agrocybe 4
aegerita (AA) and Hypsizigus mamoreus (HM) were obtained by 100 % ammonium sulfate precipitation and a Sephacryl S-300 chromatography, and then incubated for 1 6
day at various levels (5-200 µg/mL) with human peripheral blood mononuclear cells (MNC) to prepare conditioned media (CM) (AA- or HM-MNC-CM). It was observed that 8
MNC-CM prepared with AA and HM proteins at levels of 5-50 µg/mL exhibited a dose-response growth inhibition of 45-90 % on human leukemic U937 cells, while 10
unapparent results were determined when cells were cultured in the presence of proteins at the corresponding level. It suggests that activation of MNC is responsible for the 12
antiproliferation of tumor cells. In addition, assays for cytoplasmic superoxide production by the differentiated U937 cells induced by 25 µg/mL AA and HM proteins also indicated 14
a 53 and 66 % NBT (nitroblue tetrazolium) positive percentage, respectively. Furthermore, the percentage of immature blast cells decreased with the increasing level (5-25 µg/mL) 16
of proteins used to prepare MNC-CM, suggesting the potent differentiation-inducing effect of AA- and HM-MNC-CM on U937 cells. Phagocytotic activity (%) of differentiated 18
myeloid U937 cells increased in a dose-response manner and reached a value of about 33 % when U937 cells were incubated with 25 and 50 µg/mL proteins from AA. Assays 20
for the expressions of monocyte-associated antigens, CD11b, CD14 and CD68, indicated that 50-70 % of U937 cells were differentiated into monocytes/macrophages when they 2
were incubated with MNC-CM prepared with 50 µg/mL AA proteins (AA-MNC-CM-50) and HM proteins (HM-MNC-CM-50). High levels of interleukine (IL)-1β (1120 pg/mL) and 4
tumor necrosis factor (TNF)-α (1235 pg/mL) were detected in AA-MNC-CM-50, revealing the potent differentiation-inducing and anti-tumor activities of MNC-CM.
6
Granulocyte-macrophage colony-stimulating factor (GM-CSF) concentration in AA-MNC-CM-50 was detected to be 320 pg/mL using a solid-phase enzyme-linked 8
immunosorbent assay (ELISA). 10
Keywords: Anti-proliferation; differentiation; leukemic U937; Agrocybe aegerita; Hypsizigus mamoreus
1. Introduction
Dietary mushrooms contain some bioactive components such as terpenoids, 2
steroids, phenols, nucleotides, polysaccharides and proteins to present anti-tumor and immunomodulatory, antiviral and antibacterial actions (Jong and Birmingham, 1993; Ko et 4
al., 1995; Wasser and Weis, 1999). Ikekawa et al. (1982) indicated that polysaccharide (PS) fractions, EA3 and EA5, from Flammulina velutipes Sing were potent antitumor 6
agents. Besides, proflamin, a glycoprotein with a molecular weight of 1.3 ±0.4 kDa, isolated from above mushroom was orally administrated at a dosage of 10 mg/Kg bw to 8
extend the lifespan of B-16 mice implanted with Sarcoma S-180 by 85 % (Ikekawa et al., 1985). Mice fed with soluble glucan from Pleurotus ostreatus for one week improved the 10
phagocytotic activity of blood monocytes, and thus, this mushroom was considered to be one of the potential immunoenhancing materials (Paulik et al., 1996). In addition, lectin 12
from P. ostreatus exhibited strong growth inhibition on sarcoma S-180 and Hepatoma H-22 by 88 % and 75 %, respectively (Wang et al., 2000). Therefore, investigations and 14
development on the antitumor activity of dietary mushrooms as one of alternative cancer therapies might be an effective strategy in the treatment of cancer patients.
16
Agrocybe aegerita (AA) and Hypsizigus marmoreus (HM), belong to Bolbitiaceae and Tricholomataceae, respectively, are table mushrooms in Asian countries. Previous
18
investigations on α-(1→3)-D-glucan from AA and β-(1→3)-D-glucan from HM showed suppressive activity on sarcoma S-180 implanted in mice (Kiho et al., 1989; Ikekawa et 20
al., 1992). The indole derivatives in methanol extracts from AA were potent free radical scavenging agents that reduced the oxidation of fats in rat liver microsomes (Kim et al., 2
1997).
PS fraction from fruit bodies of Ganoderma lucidum was observed to be effective in 4
suppressing the proliferation and inducing the differentiation of human leukemic HL-60 and U937 cells (Wang et al., 1997). The conditioned media (CM) from PS-stimulated 6
macrophage, T lymphocytes or total mononuclear cells (MNC) contained high levels of antitumor mediators, such as interleukine (IL)-1β, tumor necrosis factor (TNF)-α and IL-6, 8
and led to apoptosis of tested tumor cells in vivo (Wang et al., 1997). Similar results were observed when MNC was treated with Cordyceps sinensis (Chen et al., 1997).
10
In the present study, in an attempt to evaluate the antiproliferation and the differentiation of U937 cells by dietary mushrooms, AA and HM, crude proteins were 12
separated to stimulate MNC to prepare CM (MNC-CM). Then, cell proliferation,
cytoplasmic superoxide production and morphological change of MNC-CM treated tumor 14
cells were determined. Assays for phagocytotic activity and expression of
monocyte-associated antigens in differentiated U937 cells were also conducted. Finally, 16
the anti-tumor mediators in MNC-CM were quantified by an enzyme-linked immunosorbent assay (ELISA).
18
2. Materials and methods
2.1. Preparation of AA and HM proteins
Fresh AA and HM form a local supermarket were blended in a cycle blender 2
(Osterizer, Mexican) with 2 parts (1:2) cold (4°C) distilled water for 1 min and then kept stirring for 24 h in a chilling room. Supernatant obtained by centrifugation (12000 × g, 4
4°C, 30 min) was treated overnight with 100 % saturated ammonium sulfate to obtain precipitates, followed by dialysis at 4°C against 100 volumes of distilled water to obtain 6
the crude proteins and by a Sephacryl S-300 gel permeation chromatography to pool the protein fractions between tube numbers 18 and 24 (column, 1.6 x 96 cm; flow rate, 42 8
ml/min; fractionation, 5 ml/tube). The thus obtained proteins were free-dried and
dissolved in PBS (phosphate-buffer saline, 8 g NaCl/1.15 g Na2HPO4/0.2 g KH2PO4/0.2 g
10
KCl/l) prior to use.
12
2.2. Preparation of MNC-CM
Human peripheral blood was obtained from each of three normal adult volunteers 14
(aged between 22-25) with informed consent, and MNC was separated by density centrifugation (400 g, 30 min) in a Ficoll-Hypaque solution (1.077 g/ml) (Pharmacia Fine 16
Chemicals, Uppsala, Sweden). A concentration of 1.5 × 106 cells/ml was incubated in 1
% glutamine (Gibco BRL, Gaitherburg, MD, USA)/10 % fetal calf serum (FCS) (Hyclone, 18
proteins in PBS to medium (5-200 µg/ml) and incubation at 37 °C in a humidified 5 % CO2 incubator for 1 day. Subsequently, the CM from AA or HM protein-stimulated MNC
2
(AA- or HM-MNC-CM) was collected by filtration through a 0.45 µm membrane and stored at –80°C until use. PHA (phytohemagglutinin) (5 µg/ml) (Sigma, St. Louis, MO, 4
USA) was also used to prepare CM (PHA-MNC-CM) and was treated as positive control. The same volume of PBS, instead of protein solution, was added to the cells to prepare 6
CM (PBS-MNC-CM). 8
2.3. Cell culture and treatments
The human leukemic U937cells, obtained from the American Type Culture 10
Collection (ATCC) (Rockville, MD, USA), were used in this study. The cells were cultured in 1 % glutamine/10 % FCS/RPMI 1640 medium and maintained in an exponential 12
growth status. Then, the cells were incubated in 35 mm petri dishes at an initial
concentration of 1 × 105 cells/ml in the presence or absence (control group) of 20 % (v/v)
14
MNC-CM (MNC-CM method). The numbers of adherent cells in day 5 cultures were collected by gently rubbing the dishes with a rubber policeman (Bellco Glass, Vineland, 16
NJ, USA) and counted using the Trypan Blue Dye exclusion test (Zweier et al., 1988) to determine the growth inhibition (%). Growth inhibition (%) = (1- cell number of MNC-CM 18
treatment/cell number of control group) × 100 %. In contrast to MNC-CM method, U937 cells incubated with purified proteins in culturing medium was termed as direct method. 20
Three separate experiments were each tested in duplicate. 2
2.4. Assay for superoxide
The production of cytoplasmic superoxide by the differentiated myeloid cells was 4
detected by the nitroblue tetrazolium (NBT) test (Lahrer et al., 1988). Cells collected from day 5 cultures were suspended in RPMI medium at a concentration of 1 × 106 cells/ml
6
and incubated at 37 °C for 30 min with an equal volume of NBT test stock solution (2 mg NBT/1 µM phorbol myristate acetate/ml PBS). The thus obtained cell suspensions (80 8
µL) were cytocentrifuged onto glass slides with the aid of a Cytospin (1200 rpm, 5 min), which were counter-stained with 0.5 % Safranin solution. The percentage of
10
formazan-containing cells was assessed out of 200 cells. Three separate experiments were each tested in duplicate.
12
2.5. Morphological examination 14
Cells collected in day 5 cultures were suspended at a concentration of 1 × 106
cells/ml in 20 % FBS/PBS and cytocentrifuged (80 µl) onto a microscope glass slides 16
with the aid of a Cytospin (1200 rpm, 5 min). Subsequently, cells were stained with Liu’s stain and observed (200 cells) under a microscope at a magnification of 1000 X. Cell 18
maturation was classified into 3 stages: immature blast cells (1), intermediates (2) and mature monocytes/macrophages (3). Three separate experiments were each tested in 20
duplicate. 2
2.6. Assay for phagocytosis
Phagocytotic activity (%) was detected according to the method described by 4
Gallagher et al. (1979). U937 cells collected in day 5 cultures were suspended in 10 % FBS/RPMI 1640 medium at a concentration of 5 × 106 cells/ml and then mixed at a ratio
6
of 1:4 with yeast suspension in PBS at the same concentration. After incubation in a humidified chamber at 37°C for 30 min, cell mixtures were placed in ice water to 8
terminate the phagocytosis and the percentage of yeast-ingesting phagocytic cells were scored out of 200 cells. Three separate experiments were each tested in duplicate. 10
2.7. Differentiating antigen determination 12
The expression of monocyte-associated antigens on the surface of U937 cells after induction of differentiation was detected according to the methods described by Goyert et 14
al. (1986) and Wang et al. (1997) with minor modifications. Cells (100 µl, 1 × 106 cells/ml)
collected from day 5 cultures were treated with 2.5 µl of primary mouse anti-human 16
CD11b, CD14 or CD68 (Serotec, Oxford, UK), and then rinsed twice with 5 % FBS/PBS, followed by reaction with 100 µl of 50-fold diluted fluorescein isocyanate
18
(FITC)-conjugated goat IgG fraction to mouse IgG (ICN, CA, USA) for 30 min in a dark and cold (4°C) place. Subsequently, 10 µl of cell suspension on a microscope glass slide 20
was examined under a light microscope (Eclipse E400, Nikon, Japan) with a
fluorescence attachment (Y-FL EPI-fluorescence attachment, Nikon, Tokyo, Japan) at an 2
excitation wavelength of 450-490 nm. CD positive percentage (%) was scored out of 100 cells. Three separate experiments were each tested in duplicate.
4
2.8. Cytokine assay 6
IL-1β, TNF-α, and GM-CSF in the various preparations of MNC-CM were quantified separately with the commercial kit (R & D, Minneapolis, MN, USA) by an ELISA at a 8
wavelength of 450 nm, according to the method described by Wang et al. (1997). The correlation coefficients (r2) for the standard curves of three cytokines were between
10
0.998-0.999. Three separate experiments were each tested in duplicate. 12
2.9. Determination of protein
Crude proteins were determined by the Bradford (1976) method using the Bio-Rad 14
protein assay dye reagent. Bovine serum albumen (0.05-0.5 mg/mL) was used to construct the standard curve (r2=0.999). Triplicate samples each were analyzed twice.
16
3. Results
18
3.1. Inhibition of U937 cell growth by MNC-CM
Figure 1 represents the changes in U937 cell growth inhibition (%) when they were 20
incubated (20 %, v/v) with various preparations of MNC-CMs. The proliferation of U937 cells was remarkably inhibited in a dose-response manner when cells were incubated 2
with MNC-CM prepared with 5-50 µg/ml AA or HM proteins. CM prepared with 5 µg/ml proteins from either HM or AA displayed about 45 % growth inhibition and that prepared 4
with 50 µg/ml proteins reached a maximal growth inhibition of 92-94 % (Fig. 1). In contrast, the presence of the corresponding level of proteins in culturing medium for 6
U937 cells (direct method) exhibited only half of the antiproliferation effect (data not shown). Further increase in proteins (50-200 µg/ml) for MNC-CM appeared to be 8
ineffective in increasing the growth inhibition. 10
3.2. NBT test
Formation of blue formazan in differentiated U937 cells was observed (data not 12
shown) and used as an indicator to differentiate from immature blast cells. NBT positive percentage increased from about 36 % to 61 % in MNC-CM prepared with 5 to 50 µg/mL 14
HM proteins, respectively, while the presence of the corresponding level of proteins in media showed relatively lower percentage in U937 cells (Table 1), suggesting that 16
activation of MNC was effective in inducing maturation of the myeloid leukemic U937 cells. Compared to those from HM proteins, AA proteins appeared to be relatively 18
stronger in inducing the leukemic U937 into differentiated cells. 20
3.3. Morphological examination of U937 cells incubated with MNC-CM
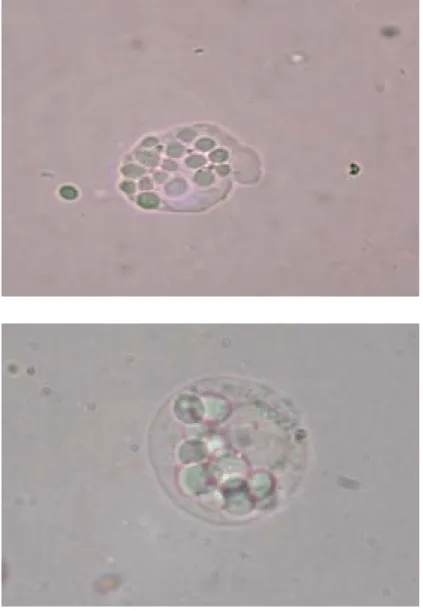
Figure 2 shows the morphological changes (magnification 1000 X) of U937 cells 2
induced by AA- or HM-MNC-CM. Immature blast cells (Fig. 2A) displayed high
nucleus/cytoplasm ratio, while maturely differentiated monocytes/macrophages showed 4
pseudopodium and vacuoles with relatively low nucleus/cytoplasm ratio (Fig. 2C). After incubation for 5 days with MNC-CM prepared with 5-50 µg/mL HM proteins, the
6
percentage of mature monocytes/macrophages increased in a dose-response manner to about 20 %, while immature blast cells dropped remarkably from about 86 % in both 8
normal and control groups to about 45 % in HM-MNC-CM-50 group (Fig. 3). No mature monocytes or macrophages were observed in normal and control groups. Compared to 10
HM proteins, AA proteins at the corresponding level appeared to display a higher percentage of differentiated cells (Fig. 3). No apparent differentiation-inducing effect on 12
U937 cells was observed when they were incubated with direct method (data not shown). 14
3.4. Phagocytotic activity of differentiated cells
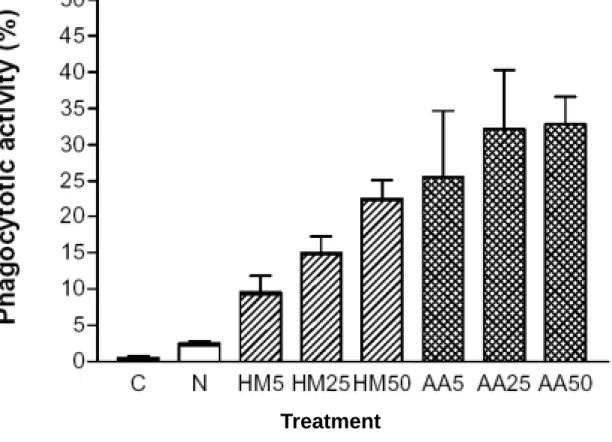
Figure 4 represents the phagocytosis of mature monocytes/macrophages, which 16
displays phagocytotic activity by ingesting of yeasts. U937 cells collected in day 5 cultures incubated with various preparations (5-50 µg/mL) of MNC-CM showed different 18
abilities to ingest foreign particles (Fig. 5). Phagocytotic activity increased with the increasing protein level in MNC-CM and reached a maximal value of about 33 % in 20
AA-MNC-CM-25 and -50 groups, revealing some certain mediators, capable of inducing differentiation of U937 cells, were released from MNC when they were stimulated with 2
mushroom proteins. However, no evident phagocytosis (0.5-2.5 %) in cells from control or normal group was observed (Fig. 5).
4
3.5. Differentiation antigen determination 6
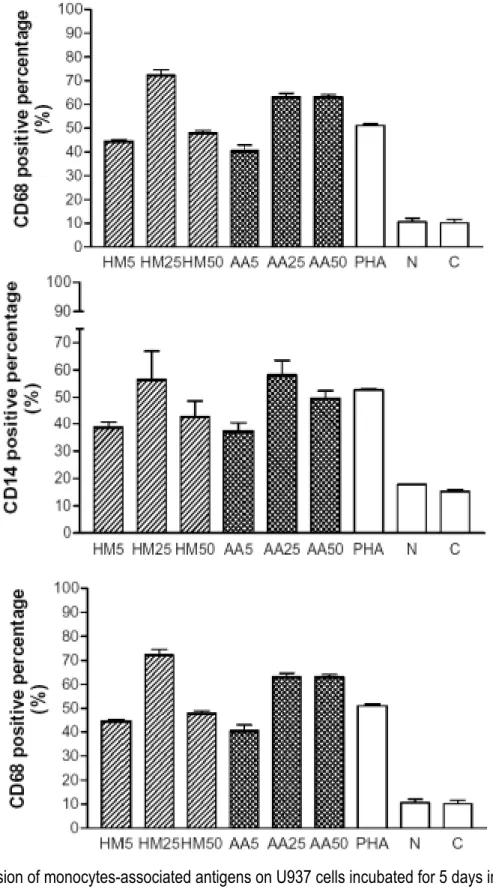
As shown in Figure 6A, CD11b positive percentage increased in a dose (5-50
µg/ml)-response manner when U937 cells were incubated with various preparations of 8
MNC-CM. Compared to 14 % in control group and 18 % in normal group, CD11b positive percentage reached 70 % in HM-MNC-CM-50 and 62 % in AA-MNC-CM-50.
10
In Fig. 6B and 6C, compared to normal group, significant (> 40 %) positive percentages of CD14 and CD 68 in the day 5 cultures of U937 cells incubated with 12
various preparations of MNC-CM were determined. PHA, a mitogen, was used to prepare MNC-CM for positive control and also displayed remarkable differentiation 14
antigen. Treatments with normal MNC-CM insignificantly induced the expressions on U937 cells of these two monocytes-associated antigens.
16
3.7. Assay for cytokines in MNC-CM 18
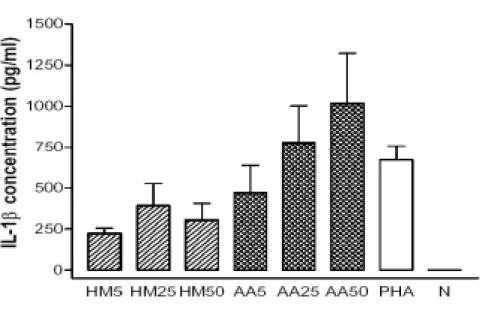
Results in Fig. 7A showed that IL-1β increased in preparations of MNC-CM in a dose (5-50 µg/ml)-response manner, and presented a level of about 800 pg/ml in 20
AA-MNC-CM-25 and 1020 pg/ml in AA-MNC-CM-50. PHA-MNC-CM (5 µg/ml), the positive control, only detected to contain 670 pg/ml. AA proteins appeared to be more 2
effective in inducing IL-1β production than HM proteins at the corresponding level. TNF-α level was detected to be 276, 720 and 1161 pg/ml in AA-MNC-CM-5, -25 4
and -50, respectively, apparently to be dependent on the dosage of stimulating proteins. Comparing to AA proteins, HM proteins appeared to be inferior to the induction of TNF-α
6
production from MNC (Fig. 7B). Results in Fig. 7C showed that GM-CSF level in
MNC-CM increased dependently on the dosage of AA and HM proteins. GM-CSF level in 8
AA-MNC-CM-50 was quantified to be 322 pg/ml, remarkably higher than that (210 pg/ml) in HM-MNC-CM-50.
10
4. Discussion
12
Mushroom PS exerts antitumor action mostly via activation of different immune responses in the host, and these bioactive components were regarded as biological 14
response modifiers (Wasser and Weis, 1999). Mizuno (1996) indicated that the possible pathways of schizophyllan, a fungus PS, against tumor were through the activation of 16
macrophages (in vitro and in vivo), which resulted in the augmentation of T-cell activities and increased sensitivity of natural killer cells to IL-2.
18
Recent reports indicated that CMs prepared from activated MNC by bioactive components from herbal or mushroom sources were effective in evaluating the immunity 20
enhancement of samples using U937 cells as tumor cell source (Wang et al., 1997; Chen et al., 1997; Liao et al., 2001; Chen and Chang, 2004). CM from PS fraction of C.
2
sinensis (10 µg/ml)-stimulated MNC markedly inhibited U937 cell growth by 78-83 % in the day 3 cultures, while the presence of the corresponding level of PS in the culturing 4
medium (direct method) displayed insignificant results. Furthermore, CM prepared with fungus PS (15 µg/ml)-stimulated MNC for 5 days exhibited a potent activity in
6
suppressing the proliferation of U937 and HL-60 cells by 87.3 and 74.7%, respectively (Chen and Chang, 2004).
8
In the present study, instead of the direct incubation of AA or HM proteins in media to interact and suppress tumor cells, various preparations (5-50 µg/ml) of MNC-CM were 10
used to observe the U937 cell growth through the activation of human peripheral blood MNC. The mode of the activation of MNC and the resulted marked antitumor effect (Fig. 12
1) were considered to be close to the immune response in host (Wang et al., 1997). Macrophages produce superoxide through the respiratory burst response as the 14
results of the reaction of oxygen with NADPH (nicotinamide-adenine dinucleotide phosphate) to form superoxide anion and the following reaction with superoxide 16
dismutase (SOD). Stronger in producing superoxide, compared to the direct method, revealed the more remarkable bactericidal effect of differentiated U937 cells induced by 18
MNC-CM method (Wang et al., 1997; Lahrer et al., 1988; Lieu et al., 1992). Similarly, PS fraction (10 µg/ml) in C. sinensis in MNC-CM was reported to be potent in differentiating 20
U937 cells by showing about 90 % NBT positive percentage (Chen et al., 1997). In addition, Liao et al. (2001) indicated that incubation with MNC-CM prepared with 400 2
µg/ml black soybean PS increased the NBT positive percentage of U937 cells to about 97 %, while insignificant NBT positive percentage was observed during the direct 4
treatment of cells. However, in contrast to MNC-CM method, presence of 5 nM bufalin in medium (direct method) led to 92 % NBT positive percentage (Zhang et al., 1991). 6
Reduction of immature blast cells of U937 cells by AA- or HM-MNC-CM was apparent, suggesting the activated MNC was capable of inducing maturation of myeloid 8
leukemic cells (Fig. 3). Similar results were observed when U937 cells were incubated with MNC-CM prepared with PS fraction from black soybean (Liao et al., 2001). Of note, 10
differentiated U937 cells was presenting phagocytotic feature of ingesting foreign particles (Figs. 4 and 5). Goliaei et al. (1998) indicated that the presence of cytokines 12
such as GM-CSF (125 pg/ml) was effective in inducing U937 cells into functional differentiated cells with 43 % phagocytotic activity.
14
Detection of differentiation antigens by ELISA was effective in evaluating the maturation of U937 cells (Chen et al., 1997). CD11b is one of the main antigen 16
expressions in monocytes, granulocytes and natural killer cells (Ault et al., 1981), while CD14 and CD68 were usually detected in monocytes and macrophages (Goyert et al., 18
1986). Results in Fig. 6 revealed that CMs from AA and HM protein-stimulated MNC contained some cytokines that helped inducing the differentiation of U937 cells and 20
resulted in the increases in the expressions of monocytes-associated antigens (Wang et al., 1997; Chen et al., 1997; Chen and Chang, 2004). Therefore, increase in these 2
antigen expressions suggested the differentiation-inducing effect of AA or HM proteins. In comparison to Fig. 6, U937 cells incubated with MNC-CM prepared with 100 µg/ml PS 4
from G. lucidum showed lower CD68 positive percentage (46.2 %) (Chen et al., 1997). IL-1β, produced by monocytes and macrophages, is a polypeptide of about 17 kDa. 6
It is capable of activating T- and B-lymphocytes and of inducing the production of GM-CSF from peripheral blood lymphocytes (Kaushansky et al., 1988). In addition, it 8
displays cytotoxicity and antiproliferation on tumors as the results of the release of TNFs and INFs by stimulating macrophages and lymphocytes (Wang et al., 1996). Results in 10
Fig. 7A were similar to the MNC-CM prepared with 100 µg/ml PS from G. lucidum (Wang et al., 1997) and to the MNC-CM prepared with 15 µg/ml PS from fu-ling (Chen and 12
Chang, 2004).
TNF-α is a polypeptide of 152 amino acids (17 kDa) produced by neutrophils, 14
activated T- and B-lymphocytes, and natural killer cells (Vilcek and Lee, 1991), and is a potent tool for tumor cell necrosis (Carswell et al., 1975). MNC-CM prepared with 100 16
µg/ml PS from G. lucidum and with 10 µg/ml PS from C. sinensis contained about 1240 pg/ml (Wang et al., 1997) and 1140 pg/ml (Chen et al., 1997) TNF-α, respectively, similar 18
to the content of this cytokine in AA-MNC-CM-50.
GM-CSF is produced by macrophages and T- and B-lymphocytes. It enhances the 20
clongenicity and differentiation of hematopoietic cells (Gasson, 1991) and U937 cells (Goliaei et al., 1998). Liao et al. (2001) assayed the GM-CSF level in MNC-CM prepared 2
with 25 and 50 µg/ml PS of black soybean to be 148 and 226 pg/ml, respectively. However, it was apparently lower than that in AA-MNC-CM-25 and -50 (Fig. 7C) in the 4
present study. The high level of GM-CSF in AA-MNC-CM suggested the strong
differentiation-inducing effect of U937 cells as shown in Table 1 and Fig. 3. In the present 6
study, insignificant level of interferon (INF)-γ in AA- or HM-MNC-CM was assayed. It could be due to the short (1 day) incubation period of time of MNC with proteins in the 8
present study. The release of INF-γ appeared to increase with the increasing incubation time for up to 5 days (Chen and Chang, 2004).
10
In summary, the release of these cytokines from activated MNC demonstrated that 12
AA and HM contain potent fungus immunomodulating proteins, which induced the production of monocytes-associated cytokines from human blood peripheral MNC, and 14
thus, led to the growth inhibition and differentiation of leukemic U937 cells. Therefore, the variety and level of antitumor mediators or differentiation-inducing factors were
16
dependent on the protein sample source. Antitumor effect of dietary AA and HM proteins was similar to that of costly fungi such as G. lucidum and C. sinensis. Their effect in vivo 18
appeared to be of interest. 20
References
Ault, K. A., Springer, T., 1981. A Cross-reaction of a rat-anti-mouse phagocyte-specific 2
monoclonal activity (anti-mac-1) with human monocytes NK cells. J. Immunol. 126, 359-364.
4
Bradford, M. M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 6
248-254.
Carswell, E. A., Old, L. J., Kassel, R. L., Green, S., Fiore, N., Williamson, B., 1975. An 8
endotoxin-induced serum factor that causes necrosis of tumors. Proc. Natl. Acad. Sci. USA 72, 3666-3670.
10
Chen, Y. J., Shiao, M. S., Lee, S. S., Wang, S. Y., 1997. Effect of Cordyceps sinensis on the proliferation and differentiation of human leukemic U937 cells. Life Sci. 60, 12
2349-2359.
Chen, Y. Y., Chang, H. M. 2004. Antiproliferative and differentiating effects of 14
polysaccharide fraction from fu-ling (Poria cocos) on human leukemic U937 and HL-60 cells. Food Chem. Toxicol. 42, 759-769.
16
Gallagher, R., Collin, S., Trujillo, J., McCredie, K., Ahearn, M., Tsai, S., Metzgar, G., Aulakh, G., Ting, R., Ruscetti, F., Gallo, R. 1979. Characterization of the continuous 18
Blood 54, 713-733.
Gasson, J. C., 1991. Molecular physiology of granulocyte-macrophage colony-stimulating 2
factor. Blood 77, 1131-1145.
Goliaei, B., Deizadji, A., 1998. Effects of hyperthermia granulocyte-marcophage 4
colony-stimulating factor on the differentiation of human leukemic cell line U937. Leuk. Res. 22, 705-710.
6
Goyert, S. M., Ferreo, E. M., Seremetis, S. V., Winchester, R. J., Silver, J., Mattison, A. C., 1986. Biochemistry expression of myelomonocytic antigens. J. Immunol.137,
8
3909-3914.
Ikekawa, T., Ikeda, Y., Yoshioka, Y., Nakanishi, K., Yokoyama, E., Yamazaki, E., 1982. 10
Studies on antitumor polysaccharides of Flammulina velutipes Sing. II. The structure of EA3 further purification of EA5. J. Pharmacobio-Dyn. 5, 576-581. 12
Ikekawa, T., Maruyama, H., Miyano, T., Okura, A., Sawasaki, Y., Naito, K., Kawamura, K., Shiratori, K., 1985. Proflamin, a new antitumor agent: preparation, physiochemical 14
properties antitumor activity. Jpn J. Cancer Res. 76, 142-148.
Ikekawa, T., Saitoh, H., Feng, W., Zhang, H., Li, L., Matsuzawa, T., 1992. Antitumor 16
activity of Hypsizigus marmoreus. I. Antitumor activity of extracts and polysaccharides. Chem. Pharm. Bull. (Tokyo) 40,1954-1957.
18
mushroom. Adv. Appl. Microbiol. 39, 153-184.
Kaushansky, K., Lin, N., Adamson, J. W., 1988. Interleukine 1 stimulates fibroblasts to 2
synthesize granulocyte-macrophage and granulocyte colony-stimulating factors. Mechanism for the hematopoietic response to inflammation. J. Clin. Invest. 81, 4
92-97.
Kiho, T., Yoshida, I., Nagai, K., Ukai, S., Hara, C., 1989. (1→3)-alpha-D-glucan from an 6
alkaline extract of Agrocybe cylindracea, and antitumor activity of its o-(carboxymethyl)ated derivatives. Carbohydr. Res. 189, 273-279. 8
Kim, W. G., Lee, I. K., Kim, J. P., Ryoo, I. J., Koshino, H., Yoo, I. D. 1997. New indole derivatives with free radical scavenging activity from Agrocybe cylindracea. J. Nat. 10
Prod. 60, 721-723.
Ko, J. L., Hsu, C. I., Lin, R. H., Kao, C. L., Lin, J. Y., 1995. A new fungal 12
immunomodulatory protein, FIP-fve isolated from the edible complete amino acid sequence. Eur. J. Biochem. 228, 244-249.
14
Lahrer, R. I., Ganz, T., Selsted, M. E., Babior, B. M., Curnutte, J. T., 1988. Neutrophils host defense. Am. Int. Med. 109, 127-142.
16
Liao, H. F., Chou, C. J., Wu, S. H., Khoo, K. H., Chen, C. F., Wang, S. Y., 2001. Isolation and characterization of an active compound from black soybean [Glycine max (L.) 18
cells. Anti-Cancer Drug 12, 841-846.
Lieu, C. W., Lee, S. S., Wang, S. Y., 1992. The effect of Ganoderma lucidum on induction 2
of differentiation in leukemic U937 cells. Anticancer Res. 12, 1211-1216.
Mizuno, T., Yeohlui, P., Kinoshita, T., Zhuang, C., Ito, H., Mayuzumi, Y., 1996. Antitumor 4
activity and chemical modification of polysaccharides from Niohshimeji mushroom, Tricholoma giganteum. Biosci. Biotechnol. Biochem. 60, 30-33.
6
Paulik, S., Mojzisova, J., Durove, A., Benisek, Z., Huska, M., 1996. The
immunomodulatory effect of the soluble fungal glucan (Pleurotus ostreatus) on 8
delayed hypersensitivity and phagocytotic ability of blood leukocytes in mice. Zentralbl. Veterinarmed. 43, 129-135.
10
Vilcek, J., Lee, T. H., 1991. Tumor necrosis factor. New insights into the molecular mechanisms of its multiple actions. J. Biol. Chem. 266, 7313-7316.
12
Wang, H. X., Ng, T. B., Ooi, V. E., Liu, W. K., Chang, S. T., 1996. A
polysaccharide-peptide complex from cultured mycelia of the mushroom Tricholoma 14
mongolicum with immunoenhancing and antitumor activities. Biochem. Cell Biol. 74, 95-100.
16
Wang, H., Gao, J., Ng, T. B., 2000. A new lectin with highly potent antihepatoma and antisarcoma activities from the oyster mushroom Pleurotus ostreatus. Biochem. 18
Wasser, S. P., Weis, A. L., 1999. Therapeutic effects of substances occurring in higher Basidiomycetes mushroom: a modern perspective. Crit. Rev. Immunol.19, 65-96. 2
Wang, S. Y., Hsu, M. L., Hsu, H. C., Tzeng, C. H., Lee, S. S., Shiao, M. S., Ho, C. K., 1997. The anti-tumor effect of Ganoderma lucidum is mediated by cytokines 4
released from activated macrophages T-lymphocytes. Int. J. Cancer 70, 699-705. Zhang, L., Nakaya, K., Yoshida, T., Kuroiwa, Y., 1991. Bufalin as a potent inducer of 6
differentiation of human myeloid leukemia cells. Biochem. Biophys. Res. Commun. 178, 686-693.
8
Zweier, J. K., Kuppusamy, P., Lutty, G. A., 1988. Measurement of endothelial cell free radical generation : evidence for a central mechanism of free radical injury in 10
2
Fig. 1. Growth inhibition (%) of U937 cells incubated with MNC-CM prepared with various levels (5-200
µg/mL) of purified proteins from Agrocybe aegerita and Hypsizigus mamoreus. U937 cells were 4
incubated in the presence of 20 % MNC-CM and vial cells were counted after 5 days of cultivation. MNC-CM was prepared after 1-day of cultivation of human peripheral blood mononuclear cells. Bars in 6
the curves represent standard error of means from three separate experiments. 8
2
4
Fig. 2. Morphologic changes of U937 cells induced by MNC-CM prepared with purified proteins from
Agrocybe aegerita and Hypsizigus mamoreus (1000 X). A: immature blast cells; B, intermediate cells; C,
6
mature monocytes/macrophage. Morphologic changes of cells were observed after 5 days of cultivation and stained with Liu’s dyes. MNC-CM was prepared after 1-day cultivation of human peripheral blood 8
mononuclear cells. Magnification × 1000. 10
A
B
2
Fig. 3. Changes in percentage (%) of differentiated U937 cells induced by 20 % MNC-CM prepared with various levels (5-50 µg/mL) of purified proteins from Agrocybe aegerita and Hypsizigus mamoreus. 4
Morphologic changes of cells were observed after 5 days of cultivation. PHA, positive group (5 µd/mL) (PHA-MNC-CM); N, normal group (PBS-MNC-CM); C, control group; HM5, 25, and 50, MNC-CM 6
prepared with 5, 25, and 50 mg HM proteins/mL, respectively; AA5, 25, and 50, MNC-CM prepared with 5, 25, and 50 mg AA proteins/mL, respectively. Bars in the columns represent standard error of means 8
from three separate experiments. 10
Fig. 4. Phagocytosis assay of differentiated U937 cells observed under microscope (Magnification 1000 X).
Treatment
2
Fig. 5. Phagocytotic activity of U937 cells incubated for 5 days with MNC-CM prepared with various 4
levels (5-50 µg/mL) of purified proteins from Agrocybe aegerita and Hypsizigus mamoreus. U937 cells were incubated in the presence of 20 % MNC-CM prepared after 1 day of cultivation of human
6
peripheral blood mononuclear cells. N, normal group (PBS-MNC-CM); C, control group; HM5, 25, and 50, MNC-CM prepared with 5, 25, and 50 mg HM proteins/mL, respectively; AA5, 25, and 50, MNC-CM 8
prepared with 5, 25, and 50 mg AA proteins/mL, respectively. Bars in the columns represent standard error of means from three separate experiments.
2
Fig. 6. Expression of monocytes-associated antigens on U937 cells incubated for 5 days in the 4
presence of 20 % MNC-CM prepared with various levels (5-50 µg/mL) of purified proteins from
Agrocybe aegerita and Hypsizigus mamoreus. Incubated U937 cells were treated with FITC-conjugated
6
goat antimouse antibody using an ELISA method. PHA, positive group (5 µg/mL) (PHA-MNC-CM); N, normal group (PBS-MNC-CM); C, control group; HM5, 25, and 50, MNC-CM prepared with 5, 25, and 8
50 mg HM proteins/mL, respectively; AA5, 25, and 50, MNC-CM prepared with 5, 25, and 50 mg AA proteins/mL, respectively. Bars in the columns represent standard error of means from three separate 10
2
Fig. 7. IL-1β, TNF-α and GM-CSF contents in MNC-CM prepared with various (5-50 µg/mL) levels of 4
purified proteins from Agrocybe aegerita and Hypsizigus mamoreus to incubate with human peripheral blood mononuclear cells for 1 day. PHA, positive group (5 µg/mL) (PHA-MNC-CM); N, normal group 6
(PBS-MNC-CM); HM5, 25, and 50, MNC-CM prepared with 5, 25, and 50 mg HM proteins/mL, respectively; AA5, 25, and 50, MNC-CM prepared with 5, 25, and 50 mg AA proteins/mL, respectively. 8
Table 1
Comparison of the NBT positive percentages of U937 cells cultured with MNC-CM methoda or direct
2
methodb using purified proteins from Agrocybe aegerita and Hypsizigus mamoreus
4
--- Proteins source Concentration NBT positive percentage (%) 6
(µg/mL) MNC-CM methoda Direct methodb
--- 8 5 35.8 ± 6.1 4.5 ± 0.6 Hypsizigus mamoreus 25 53.0 ± 6.4 12.5 ± 0.8 10 50 60.7 ± 2.9 15.6 ± 0.7 --- 12 5 52.9 ± 0.2 3.0 ± 0.5 Agrocybe aegerita 25 65.7 ± 1.7 8.7 ± 0.7 14 50 70.4 ± 3.3 10.2 ± 0.9 --- 16 PHA 5 44.7 ± 2.3 2.5 ± 0.6 Normal 4.2 ± 1.7 2.1 ± 0.5 18 Control 0.8 ± 0.3 1.0 ± 0.2 --- 20
Results from three separate experiments are expressed as mean ± SEM.
aU937 cells were incubated for 5 days in the presence of 20 % MNC-CM, which was prepared after 1
22
day of cultivation of human peripheral blood mononuclear cells with mushroom proteins
bU937 cells were incubated for 5 day in 10 % FBS/RPMI 1640 medium added with mushroom proteins
24
PHA, positive group (5 µg phytohemagglutinin/mL) Normal, PBS MNC-CM group
26