國立臺灣大學生命科學院漁業科學研究所 碩士論文
Institute of Fishery Science College of Life Science
National Taiwan University Master thesis
斑石鯛人工孵化繁殖與飼料中不同含量蛋白質對斑石鯛幼魚 成長與體組成之影響
Artificial Breeding, Rearing and Dietary Protein Levels on Juvenile Growth of Spotted Knifejaw, Oplegnathus punctatus
林宗翰 Tsung-Han Lin 指導教授: 廖文亮 博士
冉繁華 博士 Advisor: Wen-Liang Liao, Ph.D,
Fan-Hua Nan, Ph.D.
中華民國 106 年 7 月
i
謝辭
本論文承蒙指導教授 廖文亮博士、冉繁華博士於研究期間的指導,使學生 於課業知識及為人處事方面受益良多,並於論文撰寫期間之批閱及斧正,使本 論文得以順利完成,在此獻上最深的謝意。
口試期間承蒙海洋大學 沈士新博士、台灣大學 王永松博士於百忙之中細 心校閱,不吝指正並惠賜卓見,使本論文更趨完備,特此致上最深謝意。
感謝涂智欽技士,自大二至今教導我許多活生物養殖知識並提供珍貴建議,
使實驗順利進行,如父親般之照顧讓我找回學習方向;感謝漁科所同學漢唐、
紹葳、博婷、郁閔於課堂生活上之交流與照顧;感謝學長政廷、宗樺、義峰在 研究期間之陪伴以及實驗的鼎力相助,一起交流專業知識、度過低潮與失敗,
謝謝你們對我的包容,榮幸有你們陪伴;感謝佳瑜學姊於研究期間之照顧與論 文修正,希冀未來可以與各位一起精進養殖實務,共創美好未來;感謝養殖場 幹部譽高、宇聰、坤諳、俊維、傑元、韋軒、舒甯、祖賢於養殖生物照顧及實 驗操作之配合協助,彼此互相照應與養殖技術切磋,使我實驗順利完成。
最後感謝大學期間結識的大陸交換生暨好友一晗在我低潮時拉我一把,讓 我有機會獲得更多學識資訊及人生哲理;謝謝兄弟藝臻和我一起分享生活點滴、
人生規畫之建議、一起同甘共苦;謝謝土豪可望、白芍麗芳等友人在台灣及大 陸期間的交流與相助,感謝你們的一路陪伴與支持,讓我在人生低潮時找到自 己的核心價值、生活目標與自信心,願你們一同分享我的喜悅與成就。
iii
中文摘要
本實驗分為二個部分: 實驗一為開發斑石鯛人工孵化繁殖技術。實 驗二為飼料中添加不同蛋白質對斑石鯛幼魚成長與體組成之影響。
實驗一對斑石鯛種魚藉由人工催熟使其性腺成熟,並培育斑石鯛,
建立斑石鯛胚胎發育、仔稚魚形態發育及人工孵化繁殖餌料生物投餵 序列。生殖季期間以塑膠軟管檢視種魚成熟度後,施打 HCG 1000 I.U.
/ kg、LHRH - A2 20 μg / kg 促進其成熟後於傍晚昏暗時將斑石鯛雌魚 取出擠卵後,再以斑石鯛精子進行人工授精,平均受精率為 71.20 ± 0.10 %,受精卵為淡黃色、單一油球之浮性透明卵,受精卵平均直徑為 1.03 ± 0.04 mm,油球徑為 0.23 ± 0.01 mm。水溫 23 ± 1 ℃ 下,受精 後 3 hr 40 min 進入桑椹期;受精後 7 hr 50 min 進入原腸期;受精後
14 hr 25 min 進入神經胚期,受精後 15 hr 10 min 進入器官形成期;受 精後 29 hr 30 min 胚胎孵化,剛孵出仔魚體標準長 1.96 ± 0.01 mm,具 一橢圓卵黃囊,長徑 1.05 ± 0.01 mm;短徑 0.65 ± 0.01 mm;油球徑 0.21
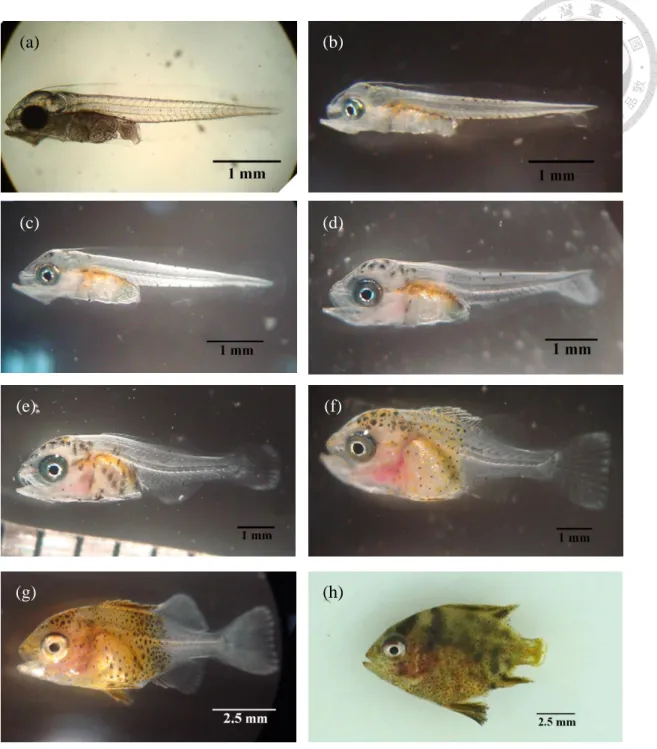
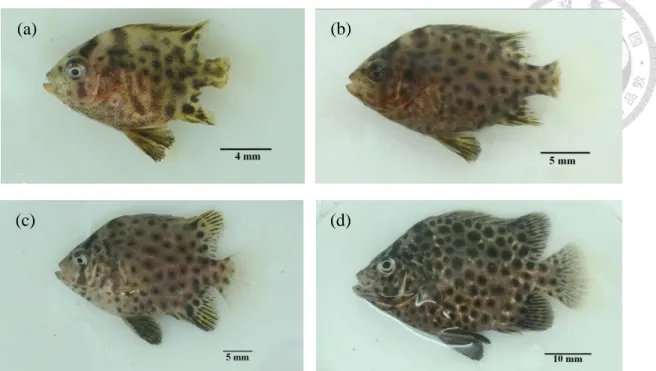
± 0.01 mm,孵化後第 3 天卵黃囊完全吸收,魚體標準長 3.65 ± 0.05 mm。仔魚一開口可攝食輪蟲與橈腳類無節幼蟲,隨成長投餵較大體型 之橈腳類幼生及成蟲,孵化後第 21 天進入稚魚期,第 23 天後增加 餵食豐年蝦無節幼蟲,第 28 天除餵食餌料生物並開始以少量商業飼 料馴餌,至第 44 天進入幼魚期,魚體標準長 25.98 ± 3.70 mm,黑色
圓點數量增加,體色由乳白色轉為黑褐色,此時已可完全投餵商業人 工飼料。斑石鯛仔魚與稚幼魚發育在水溫 26 ± 1 ℃ 下,第 44 天完 成變態。
實驗二以紅魚粉為蛋白質來源,鱈魚肝油與玉米油為油脂來源,
配製等油脂 (7 %) 等能量之六組不同蛋白質 (35 %、40 %、45 %、50 %、
55 %、60 %) 飼料投餵平均初重約 16 g 斑石鯛幼魚 8 週。結果顯示 水溫 29 ± 1 ℃ 下,斑石鯛幼魚增重率在 227.79 ± 46.2 % - 477.47 ± 24.42 % 之間,SGR 在 2.10 ± 0.26 - 3.13 ± 0.08 之間,FCR 在 1.05 ± 0.13 - 1.67 ± 0.49 之間,各成長指標隨著飼料蛋白質含量增加而增加,
並在蛋白質含量 55 % 組有最高增重率 (477.47 ± 24.42 %),與蛋白質 含量 35 % - 50 % 組有顯著差異 ( p < 0.05),折線回歸求得最適蛋白質 含量為 57.58 %。增重率、飼料轉換率及特殊成長率在蛋白質含量 35 % - 55 % 時會隨著蛋白質含量增加而改善。蛋白質效率 (PER) 在 1.61 ± 0.24 - 1.91 ± 0.38 之間,隨著飼料蛋白質含量上升而下降,並在蛋白質 含量 60 % 組為最低 (1.61 ± 0.24 ),各組之活存率最佳為 96 %,飼料 蛋白質含量 35 % 組別之活存率為 36 %,低於飼料蛋白質含量 40 % - 60 % 組別。
關鍵詞:斑石鯛、人工孵化繁殖、胚胎發育、仔稚魚發育、蛋白質需求
v
Abstract
This study aims to promote artificial rearing and breeding techniques of Oplegnathus punctatus via record of embryonic ontogeny, larval and juvenile development and establishment of a feeding program for the species, and then assigned to experiment II to know the effect of dietary protein level on the growth and body composition of O. punctatus juvenile.
In the first trial, used the injection of hCG 1000 IU/kg BW and LHRH - α 20μg / kg treatment to induce spawning of female O. punctatus and were artificially fertilized from April to May
2017. The fertilized pelagic eggs are transparent with diameter of 1.03 ± 0.04 mm and single oil globule with diameter of 0.23 ± 0.01 mm. The eggs hatched in 30 hours after fertilization at 23 ± 1
℃. The newly-hatched larvae are 1.96 ± 0.01 mm in standard length. From 3 to 33 dph the larvae can be fed with rotifers and copepods enriched with Nannochloropsis oculata and Tetraselmis chui, bigger copepods and nauplii can be fed as larvae grows up.
Metamorphosis occurred in 28 dph and artificial feed was mixed
with artemia to train the larvae to accept. After 44 dph, the larvae
developed into juvenile stage with 26 mm in standard length.
In the second trial, O. punctatus juvenile with initial weight 16 g fed on six diets with same energy and lipid level (7 %) but different dietary protein levels of 35, 40, 45, 50 %, 55 % and 60
% for eight weeks. The result showed the weight gain were between 227.79 ± 46.24 % - 477.47 ± 24.42 %, specific growth rate (SGR) were between 2.10 ± 0.26 - 3.13 ± 0.08, feed conversion ratio (FCR) were between 1.05 ± 0.13 - 1.67 ± 0.49.
Fish fed diet with 55 % protein has the highest weight gain (477.47 ± 24.42 %), and was significant higher than diets with 35 - 50 % protein (p< 0.05). Based on the weight gain using broken-line model analysis showed the optimal dietary protein level for O. punctatus juvenile with initial weight of 16 g cultured under 29 ± 1 ℃ is 57.58 %. Weight gain, SGR were between 2.10 ± 0.26 - 3.13 ± 0.08 and were improved with dietary protein increased from 35 % to 55 %. SGR in diets with 55 % protein was significant higher than diets with 35 - 50 % protein (p< 0.05).
FCR were between 1.05 ± 0.13 - 1.67 ± 0.49 and improved with
dietary protein increased. Fish fed 55 % has lowest FCR (1.05 ±
vii
0.13) and significant lower than groups fed 35 - 45 % protein diets. Coefficient factor (CF) was not significantly increased by dietary protein level among 40 - 60 % groups (p> 0.05).
Hepatosomatic index (HSI) and viscerosomatic index (VSI) was not generally affected with dietary protein. Crude protein in muscle increased with increasing dietary protein. Crude lipid and Ash in muscle was not significantly affected with dietary protein among 40 - 60 % groups.
Key words: Oplegnathus punctatus, spotted knifejaw, artificial
breeding, embryonic development, larval development, dietary
protein requirement.
目錄
口試委員審定書 ... i
謝辭 ... ii
中文摘要 ... iii
英文摘要 ... v
目錄 ... viii
表目錄 ... ix
圖目錄 ... xi
前言 ... 1
文獻整理 ... 3
材料方法 ... 27
結果 ... 38
討論 ... 50
結論 ... 72
參考文獻 ... 73
ix
表目錄
Table 1. The optimal protein level and protein source of fish. ... 103
Table 2. Composition of experimental diets for juvenile spotted knifejaw of experiment 2. ... 108
Table 3. Embryonic development of Oplegnathus punctatus from fertilization to newly hatched larvae at 23 ± 1 ℃. ... 109
Table 4. The initial, final weight, weight gain, FCR, SGR, FI, PER (mean ± S.D.) and survival of Oplegnathus punctatus of experiment 2 in 8 weeks.
... 111
Table 5. CF, VSI and HSI of spotted knifejaw (Oplegnathus punctatus) of experiment 2 in 8 weeks. ... 112
Table 6. Proximate analysis of muscle of spotted knifejaw (Oplegnathus punctatus) of experiment 2 in 8 weeks. ... 113
圖目錄
Figure 1. Broodstock of spotted knifejaw (Oplegnathus punctatus). .... 114
Figure 2. Oocyte of spotted knifejaw (Oplegnathus punctatus) before hormonal treatment. ... 114
Figure 3. Morphological changes of female spotted knifejaw (Oplegnathus punctatus) after hormonal treatment. ... 115
Figure 4. Cloaca of mature female spotted knifejaw (Oplegnathus punctatus) before spawning. ... 115
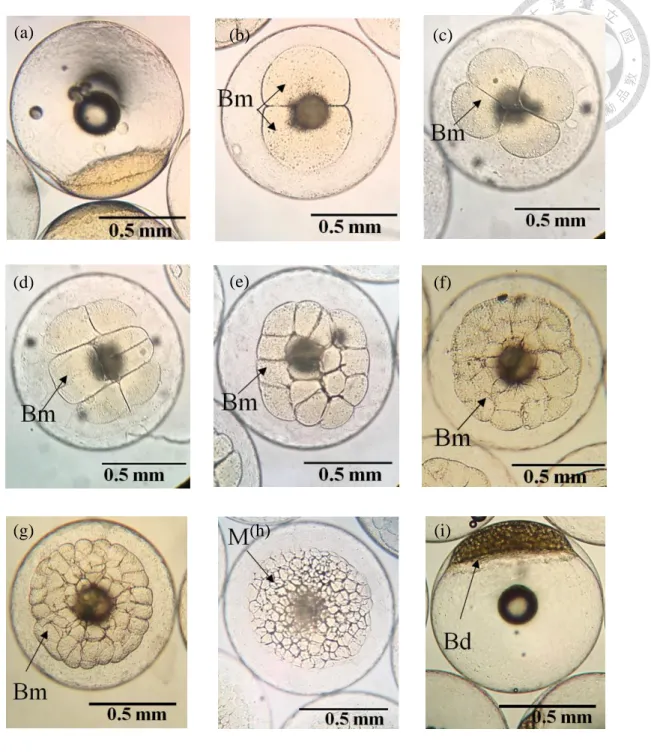
Figure 5. Embryonic development of Oplegnathus punctatus from fertilization to high blastula stage at 23 ± 1 ℃. ... 116
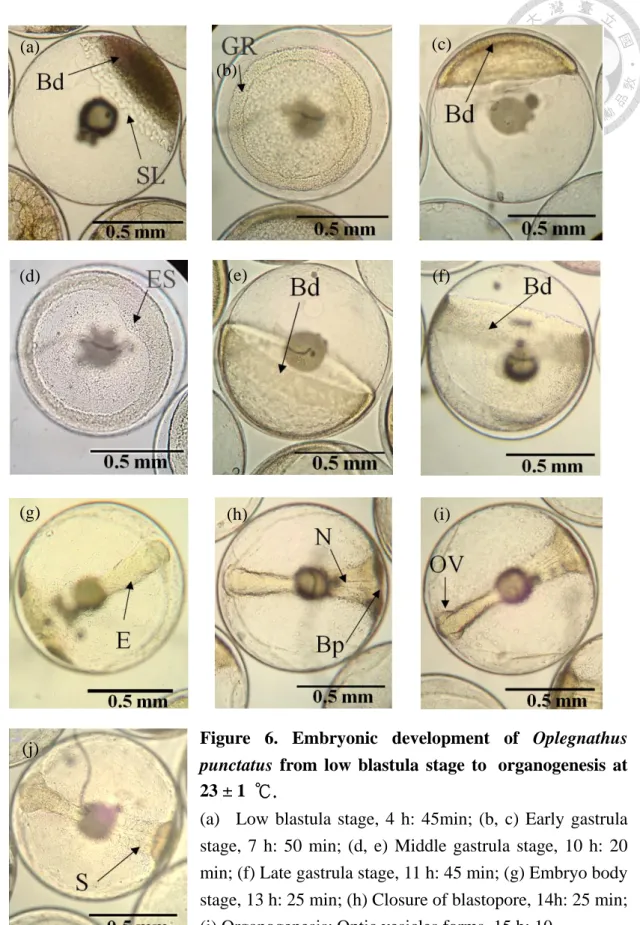
Figure 6. Embryonic development of Oplegnathus punctatus from low blastula stage to organogenesis at 23 ± 1 ℃. ... 117
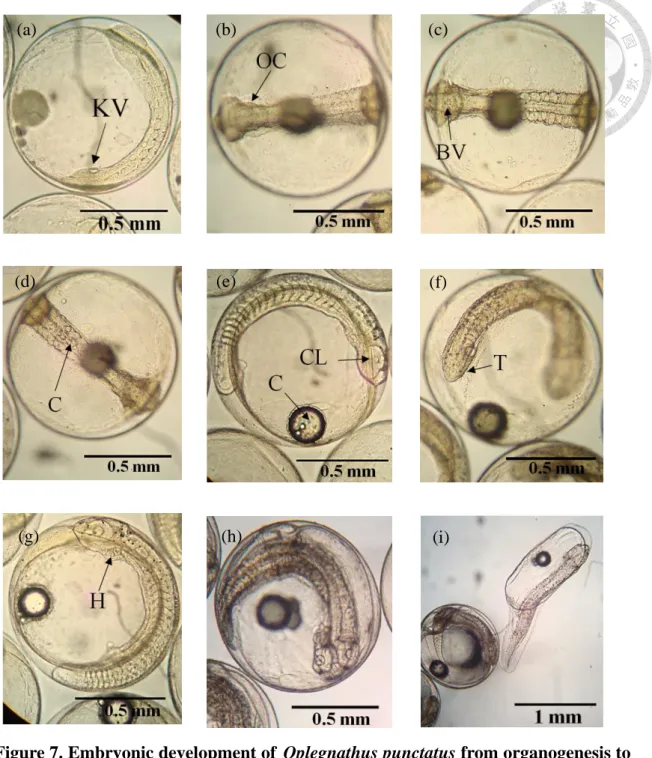
Figure 7. Embryonic development of Oplegnathus punctatus from organogenesis to Pre-hatching stage at 23 ± 1 ℃. ... 118
Figure 8. The morphological changes of Oplegnathus punctatus in larval
xi
Figure 9. The morphological changes of Oplegnathus punctatus from larval stage stage to Metamorphosis stage at 26 ± 1 ℃. ... 120
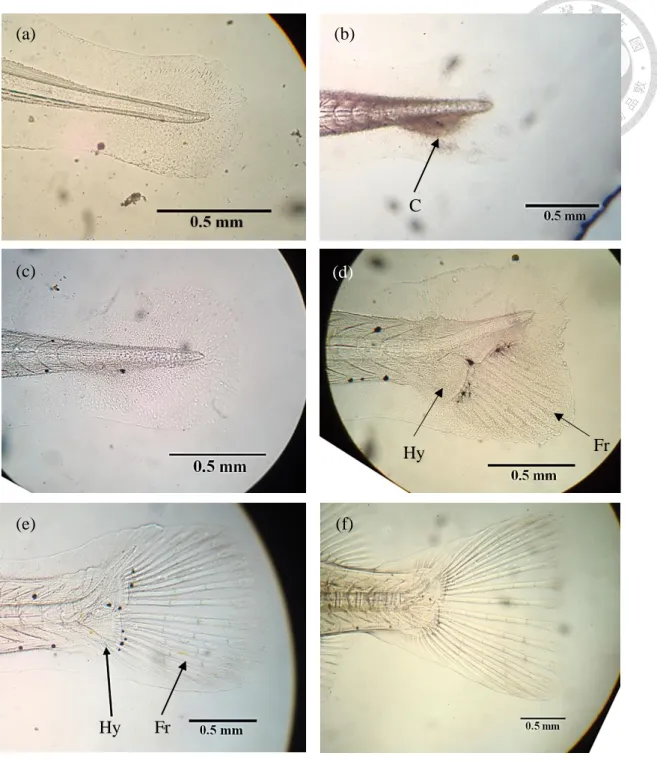
Figure 10. The morphological changes of Oplegnathus punctatus from Metamorphosis stage to juvenile stage at 26 ± 1 ℃. ... 121 Figure 11. The development of caudal fin skeletons in Oplegnathus punctatus from 2 dph to 21 dph stage at 26 ± 1 ℃. ... 122
Figure 12. Development of vertebral column, dorsal fin and anal fin in Oplegnathus punctatus from 11 dph to 21 dph stage at 26 ± 1 ℃. ... 123
Figure 13. Feeding program and growth of Oplegnathus punctatus at 26 ± 1
℃ from 0 to 54 days post-hatch. ... 124
Figure 14. Broken - line regression of weight gain (y) on dietary protein level (x) of juvenile Oplegnathus punctatus (initial TW > 16 g) in experiment II in 8 weeks ... 125
前言
斑石鯛 (Oplegnathus punctatus) 屬於條鰭魚綱 (Actinopterygii)、
新鰭魚亞綱 (Neopterygii)、鱸形目 (Perciformes)、鱸亞目 (Percoidei)、
石鯛科 (Oplegnathidae)、石鯛屬 (Oplegnathus) 的魚類 (沈,1993),
全世界共有七種石鯛。台灣物種名錄資料顯示,台灣海域所產的石鯛 屬只有條石鯛 (O. fasciatus) 與斑石鯛 (O. punctatus) 兩種。石鯛主要 分布於太平洋和印度洋沿岸,包括台灣、日本、韓國、中國、夏威夷 等熱帶、亞熱帶的沿近海礁岩岸及珊瑚礁區 (林等,2013)。斑石鯛,
牙齒尖利,可以咬碎貝類、龍蝦、海膽等的堅硬外殼,食用價值極高,
肉質細膩,口感獨特,富含多種氨基酸與不飽和脂肪酸,在日本料理 中具有「刺身絕品」之譽 (尤等,2015)。大陸市場上流通的斑石鯛苗 8
cm 可達 16 元人民幣,每公斤售價約 600 元人民幣以上,屬高經濟 價值魚種,因此建立斑石鯛人工繁養殖及魚苗培育技術則相當重要。
全球氣候變遷影響下,澎湖 2008 及 2011 年冬天,出現歷年來少見 的淺海寒災,受到冬季強勁的東北季風及持續性的低溫海水影響,不 管是澎湖海域的野生魚類,或是箱網養殖魚類都傳出大量凍死的災情。
兩次寒害凍死的魚類多達 64 科 183 種 (呂,2012;Hsieh et al., 2008),
其中未見石鯛科魚種,可知石鯛可耐受澎湖冬季的低溫海水,因此石 鯛有潛力成為未來海洋箱網養殖的新興對象魚種之一 (林等,2013)。
2
水產養殖業最重要的成本主要有二項,分別是飼料成本及種苗成本 (鄭,
2006),其中最昂貴的主要成分為蛋白質 (Shiau and Lan, 1996),其中魚 粉為水產飼料之主要成分,為飼料中動物性蛋白質的主要來源,其價 格高低都直接影響生產成本。現今因氣候變遷導致國際魚粉價格不段 攀升,飼料成本提高而獲利減少,飼料配方中首重蛋白質含量,若飼 料蛋白質過低將造成水生生物生長下降,含量過高則使生物將過多蛋 白質作為能量消耗而無法合成體蛋白造成蛋白質利用上的浪費。因此 本研究分為二個部分: 實驗一探討斑石鯛人工孵化繁殖技術,觀察其胚 胎發育仔稚魚之發育過程並建立餌料生物投餵次序,並將培育之幼苗 育成。實驗二以魚粉做為主要飼料蛋白來源,配置不同蛋白質含量,
等油脂等能量的飼料投餵斑石鯛,探討其幼苗成長率及體組成的影響,
以推估飼料最適蛋白質添加量。
文獻整理
一、斑石鯛 (Oplegnathus punctatus) 之簡介
1. 斑石鯛的分類及形態特徵
輻鰭魚綱 Class Actinopterygii
新鰭魚亞綱 Subclass Neopterygii 鱸形目 Order Perciformes
鱸亞目 Suborder Percoidei
石鯛科 Family Oplegnathidae 石鯛屬 Genus Oplegnathus
斑石鯛 Oplegnathus punctatus
(沈,1993) 斑石鯛,又俗稱斑鯛、黑金鼓 (張等,2015),英文名為 Spotted
knifejaw。斑石鯛外型延長呈橢圓形,側扁而高。頭小,前端鈍尖。吻 短,眼大側位。頜齒癒合成鸚鵡喙嘴狀,鋤骨和齶骨均無齒。前鰓蓋 骨後緣具鋸齒;鰓蓋骨上具 1 扁小棘。體被細小櫛鱗,吻部、眼間隔 及頰部無鱗,各鰭基底均被小鱗;背鰭單一,硬棘 XII,軟條數 17;
臀鰭硬棘 III,軟條數 12 - 13;尾鰭截形。體灰褐色,有銀白光澤。頭 部、體側、胸鰭及各奇鰭基部分佈許多黑斑,幼魚的體色較淡呈褐色;
各鰭暗色,最大體長 86 cm (沈,1993)。斑石鯛肉質細膩,口感獨特,
4
富含多種氨基酸與不飽和脂肪酸,食用價值極高,在日本料理中具有 刺身絕品之稱 (尤等,2015)。文獻指出,斑石鯛肌肉中多不飽和脂肪 酸百分比含量為 35.24 %,DHA 和 EPA 含量分別為 12.4 %和 3.72 %;
肌肉中礦物質元素 Ca,Mg 和 P 的含量分別為 720,1510 μg/g 和 9930 μg/g,微量元素中鋅的含量最高(15 μg/g),其次為鐵(4.9 μg/g)。斑
石鯛肌肉中富含必需氨基酸且 DHA 和 EPA 及礦物質元素含量豐富,
是營養價值較高的優質食用魚類 (王等,2016)。
2.地理分佈
斑石鯛分佈於太平洋區,包括夏威夷群島、關島、澳洲、菲律賓、
臺灣、日本,及中國大陸沿海等。臺灣北部、東部、西部、南部及離 島之岩礁,珊湖礁區皆產。模式種產地位於日本長崎 (沈,1993)。
3. 生態習性
斑石鯛為溫熱帶沿近海魚類,棲於近海珊瑚岩礁 20 - 100 m 海底 區域,幼魚隨著海藻漂移。斑石鯛為肉食性,齒銳利,可咬碎貝類、
龍蝦或海膽等堅硬之外殼 (沈,1993;尤等,2015),因此又俗稱海膽 鯛、黑嘴、硬殼仔 (林等, 2013),斑石鯛在天然海域中全年皆產,主 要漁法為底拖網、延繩釣及手釣,全年皆產,無明顯盛漁期。台灣南 部地區較多,產量不大 (沈,1993)。
4. 生長特性
斑石鯛生存水溫為 6 - 32 ℃,適宜生長水溫為 22 - 28 ℃,其生 命力強,生長速度快、抗逆性強,適於箱網、池塘及工廠化等養殖環 境 (尤等,2015)。文獻指出,以中國海南省三亞地區養殖斑石鯛為例,
斑石鯛經過半年飼養體重可達 500 g (張等,2015)。另有文獻指出,日 齡 68 天的斑石鯛稚魚 (全長 5.3 ± 0.3 cm,體重 3.6 ± 0.7 g), 在平 均水溫 25.3 ℃ 條件下,投餵配合飼料,經過 254 天的養殖,平均體 重可達 659 g,養殖活存率 94 %,餌料係數 1.67 (川辺與木村, 2007b),
斑石鯛生長迅速,初重 3.6 g 種苗經過 10 - 12 個月養殖,體重可達 450 - 500 g (尤,2015)。
5. 生殖特性
石鯛為雌雄異體 (區等,2014),5 - 6 齡魚即可達成熟體型,在全 長 43.4 - 59.1 cm,體重 1.9 - 5 kg 即可成功自然產卵,生殖期介於 4 月底至 6 月底,水溫 21.1 - 24.6 ℃ 之春末夏初之際 (川辺與木村,
2007a)。斑石鯛性成熟時,雄魚身體斑紋消失體色轉為黑色,並出現白 斑;雌性個體身體斑紋明顯且身體轉為灰褐色,且二性別成熟者嘴喙 皆轉白色。雄魚成熟體型較雌魚大 (道津與夏苅, 1967)。藉由種魚餌料 添加大豆卵磷脂、磷蝦、冷凍魷魚、竹筴魚及綜合維生素進行營養強 化即可使之自然產卵 (川辺與木村, 2007a)。藉由延長光照週期及人工 升溫處理可將其生殖季提前至 2 個月 (熊井,2005)。
6
6. 斑石鯛幼苗培育
王 (2015) 報告指出,斑石鯛受精卵於水溫 22 ± 1 ℃ 及海水鹽度 29 ± 0.5 ppt 的孵化條件下,經過 32 hr 左右孵化。孵化的幼苗全長 3.49 ± 0.12 mm,至第三天魚苗全長 3.81 ± 0.12 mm,此時卵黃囊消失,
消化道形成,肛門與外界接通;上下頜開啟,仔魚開口並攝食輪蟲,
第四天魚鰾開始發育,17 dph (days post hatch) 稚魚全長 5.66 ± 0.21
mm,此時在投餵餌料時,其搶食行為增加。消化能力加強,開始由輪 蟲向豐年蝦、橈腳類等生物餌料過渡,50 dph 幼魚全長 43.00 ± 0.52
mm,此時的幼魚體型接近成魚,幼魚體被櫛鱗,與成魚型態一致,文 獻指出,以海洋擬球藻、濃縮小球藻、β-胡蘿蔔素及酵母二次培養後 之輪蟲投餵斑石鯛幼苗可改善其孵苗後期 (15 dph) 活存率低的問題
(藤田等, 1999;織田等, 2000;高野瀬等, 1999, 2000, 2002),川辺與木 村 (2007b) 指出,斑石鯛後期培育階段,尚須適當調整幼苗蓄養密度 以及餌料營養強化,可有效降低幼苗殘食以及營養不良現象。
7. 斑石鯛養殖現狀
斑石鯛之自然產卵由日本近畿大學首次成功,並指出斑石鯛生長 速度較條石鯛快 (原田等, 1979)。此後於鹿兒島縣栽培漁業中心藉由生 餌投餵斑石鯛種魚,每年皆可採集大量受精卵做為孵化繁殖 (富安等, 1994, 1995)。宮下 (2000) 指出,斑石鯛具備: (1). 成長速度快 (2). 市
場價格昂貴 (3). 飼料效率佳等優點,因此未來推廣斑石鯛之人工繁養 殖極具市場潛力,石橋 (2000) 指出,斑石鯛除了可作為單養物種,亦 可與鰤魚 (Seriola quinqueradiata) 作為混養對象。然而,斑石鯛養殖 過程中有諸多瓶頸待克服,文獻指出,斑石鯛幼苗養殖過程容易受貝 尼登吸蟲 (Neobenedenia Girellae) 、海水白點蟲 (Cryptocaryon irritans) 及卵圓鞭毛蟲 (Amyloodinium ocellatum) 感染 (張等,2015;范,2016;
川辺與木村, 2007b)。當飼養水溫超過 23 ℃時,斑石鯛幼苗容易感染 上皮囊腫症 (Epitheliocystis disease),另外對神經壞死病毒 (NNV)、斑 石鯛虹彩病毒 (SKIV) 具感染性 (中村等, 1988;高野瀬等, 2001;Egusa et al., 1987; Dong et al., 2010)。因斑石鯛生長快速,養殖過程需針對不
同階段調整蓄養密度,其中過高蓄養密度容易造成斑石鯛魚苗生長停 滯,並且因個體間成長速率差異而發生殘食 (愛媛県水産試験場,1994;
川辺與木村, 2007a,b),富安等人 (1994) 指出,斑石鯛幼苗培育時期容 易因餌料營養不良造成幼苗畸形,近年日本針對斑石鯛繁養殖之飼育 水槽、受精卵採集、受精卵消毒、飼育水溫、種魚及幼苗餌料之投餵 流程、幼苗中間育成皆已詳細紀載 (原田等, 1979;川辺與木村, 2007a,b)。
中國於 2014 年首次於山東萊州明波水產有限公司藉由人工調控促進 斑石鯛種魚性腺成熟,使其排卵受精且成功培育幼苗,並在中國南北 方推廣規模化養殖 (王,2015),更發展至中國海南省熱帶地區養殖 (張
8
等,2015),養殖方式主要有箱網養殖及室內循環水養殖 (尤等,2016)。
文獻指出,中國海南三亞地區養殖斑石鯛試驗為例,蓄養水溫高於 31
℃ 時,斑石鯛攝食性減弱且易染海水白點蟲症 (張,2015)。目前斑石 鯛研究主要為病害檢測、分子生物學、胚胎發育、換肉率與肌肉營養 成分分析及養殖模式研究 (尤等,2015;張等,2015;王等,2016;奉 等,2016;Dong et al., 2010; Shuang et al., 2013)。然而,其幼苗最適生 長營養配合飼料需求尚未有基礎研究。
四、魚類孵化繁殖
1. 種魚培育
種魚 (Broodstock) 為水產養殖產業中,以生產子代為主要用途的 成熟個體,在人工管控之蓄養環境中生產子代以取代野外採集來源,
或以放流來增加野外資源量。種魚來源有野外捕捉 (Wild caught, WC) 及人工培育 (Captive breeding, CB)。魚類由於受到產季環境等因素導致 產卵質與量不穩定,對其人工孵化繁殖是主要障礙 (Kjorsvik et al.,
1990)。野外捕捉個體對人為飼養環境會產生不適應,因此需要一至二 年的人工馴化與滋養,方可將產能有效提升,種魚繁殖表現亦與其年 齡與產齡有很大關係,Bromage et al. (1988) 指出虹鱒 (Oncorhynchus mykiss) 於第二次產季產下之發眼卵活存率相對第一次產季較高。種魚 馴養在人工環境下,光週期、水溫及微生物等因素會對受精卵品質產
生影響 (Brooks et al., 1997; Wang et al., 2010)。其中光週期與溫度是控 制溫帶地區魚類生殖週期的主要因素 (Wang et al., 2010)。飼料中的必 需脂肪酸種類與含量是影響種魚繁殖表現的因素之一,投餵生餌或種 魚專用飼料可提升種魚產能 (Watanabe et al., 1984a, b)。
2、受精卵
水生動物胚胎發育過程中以大洋浮性卵 (Pelagic egg) 為型態之一,
其具備較薄的片狀絨毛膜及均勻的卵黃,可在較短時間內孵化(Lønning et al., 1988)。Ikeda et al. (1991) 指出,黑鯛 (Acanthopagrus schlegelii)、
嘉鱲 (Pagrus major) 等魚之浮性卵,產卵後受精卵在 24 ℃ 下,約 20
hr 孵化;不同種間浮性卵外型與構造幾乎一致,難以區分種類,僅能 依據卵徑判別,或以孵化法 (Rearing method) 判斷魚花種類,浮性卵 具備略小於海水的比重,在鹽度 30 ppt 以上之海水中可漂浮於水面,
並隨潮汐及海流漂移,可有效均勻分散於環境中或遷徒至其他海域,
漂浮於水面亦可避免底棲掠食者捕食 (盧,2016)。
3.
魚類早期發育各階段劃分根據報告指出,魚類的早期發育階段可分化為胚胎期、仔魚期、
稚魚期 (變態期)、幼魚期四個階段 (徐,1984;于,2008;何,2011;
Kendall et al., 1984; Falk-Petersen, 2005)。
(1). 胚胎期 (The egg stage, embryo stage)
10
胚胎期指精卵結合為合子後,受精卵快速分裂直至仔魚孵化出膜 的發育過程。海水硬骨魚的卵多為浮性卵,卵徑大小在 0.6 - 6 mm 之 間 (于,2008),受到遺傳因素、孵化水溫、水流速度、海水鹽度等條 件影響,各種魚類的胚胎發育時間不盡相同,如牙鮃 70 hr 左右,條 石鯛 30 hr 左右,而鮭鱒類通常持續幾週,狼魚的胚胎期則需要持續 幾個月 (Pavlov,1986)。根據胚胎發育的場所以及胚胎期營養物質的 來源,魚類的生殖方式主要有卵生、卵胎生、胎生。卵生方式普遍存 在於大多數魚類中,將成熟的配子排到水中,體外受精,以卵黃囊內 存儲的有機物質作為營養來源維持胚胎發育過程;卵胎生方式多見於 鯊、鰩、魟、鱝等繁殖力較低的軟骨魚類,卵胎生的胚胎發育所需營 養物質由卵黃提供,呼吸則靠母體;胎生方式主要存在於板鰓類中,
胚胎發育過程的營養不僅依靠卵黃囊供應,同時依賴於類似哺乳動物 的胎盤狀衍生物與母體發生物質交換 (樓,1996)。依據 Kimmel et al.
(1995) 對斑馬魚胚胎發育過程的描述,一般可將魚類胚胎發育劃分為:
(1). 合子期 (Zygote period),精卵結合,形成受精卵;(2). 卵裂期 (Cleavage period),受精後短時間內合子快速分裂形成多細胞;(3). 囊 胚 期 (Blastula period) , 分 為 低 囊 胚 和 高 囊 胚 階 段 ; (4). 原 腸 期 (Gastrula period) , 發 生 原 腸 作 用 , 三 胚 層 形 成 ; (5). 器 官 形 成 期 (Segmentation period),重要功能器官原基出現;(6). 孵化期 (Hatching
period),胚胎為最後的破膜孵化做準備 (Kimmel et al., 1995)。硬骨魚 的卵母細胞在最終的成熟階段,會吸收周圍的水分,發生水合作用而 變充盈有浮性。諸如鱈魚、鮃鰈類等海水魚類,其卵細胞經過水合作 用後水分可高達 92 %, 因此卵細胞的浮性主要來自於含水量,而非 油脂類 (Craik and Harvey, 1984)。受精結束後的短暫時間內,細胞質移 向動物極一端成為胚盤,並進行盤狀卵裂。經過數次分裂之後,細胞 成團狀無法記數,並進入囊胚期,隨著發育的進行,胚盤的部分細胞 與卵黃邊緣融合,形成具有多核結構的卵黃多核層。卵黃囊在胚環下 包作用下進入體腔,在卵黃多核層的介導下,於胚胎發育過程提供能 量 (Timmermans, 1987)。原腸期是形成動物內部生理結構和外部形態 的重要階段。囊胚期之後,胚環局部增厚為胚盾,胚環沿卵黃囊下包,
形成卵黃栓,即未來的胚孔,原腸作用開始。原腸胚時期,胚胎細胞 高度重組為三胚層並形成胚體的早期結構 (Lilianna Solnica-Krezel, 2005)。
(2). 仔魚期 (The larval stage)
仔魚期是從胚體孵化出膜到鰭膜消失,各運動器官功能完善的階 段為止,此階段主要依賴浮游方式生活。根據卵黃囊是否退化,仔魚 期可分為前期仔魚 (有卵黃囊,pre-larval stage) 和後期仔魚 (卵黃囊消 失,post-larval stage)(于,2008;王,2015)。前期仔魚未開口,口、肛
12
門未與外界接通,無法攝取外源食物,只依賴卵黃囊、油球作為營養 來源供其生長發育,身體呈現透明,眼部色素較淺,前期仔魚期由於 攜帶卵黃囊,又稱之為卵黃囊期 (The yolk sac stage)(Falk-Petersen,
2005),卵黃囊期持續的時間在各種海水魚中不一致,條石鯛為孵化後 4 天 (于,2008;何 2011),條斑星鰈孵化後 7 天 (柳等,2009),大菱 鮃孵化後 4 天 (佟,2010)。仔魚階段的個體,鰓仍處於發育過程中,
需要依靠皮膚細胞與外界環境進行離子交換和氣體交流,來維持滲透 壓平衡和呼吸作用 (Peter, 2007);後期仔魚卵黃囊、油球消失,開口攝 食,仔魚開始巡遊運動。初孵仔魚的眼睛一般無色素沉著,作為視覺 性捕食者,隨著仔魚開口攝食,視杆細胞和視錐細胞功能逐漸 完善並 具有視覺功能 (O' Connell C.P., 1981; Blaxter, 1986)。剛孵化出來之仔魚 在吸收完卵黃囊內營養後,其營養來源改為向外界攝取,而這些外在 營養物質,稱為營養源 (Kohno et al., 1990; 1997)。營養源大致上分為 二大類,一種為天然餌料;水域中的天然餌料係指水生動、植物或動、
植物的屍體以及一些微生物和有機碎屑等,這些有機物質中,具生命 現象並且可當作餌料的動物或植物均統稱為餌料生物;另一種為人工 飼料: 已選擇優良的完全配方飼料替代餌料生物,提供魚苗所需的各種 營養 (鄧,2008)。
(3). 稚魚期 (The Transformation stage)
多數硬骨魚發育過程中,仔魚和幼魚之間有一個明顯的體型結構 變化的階段,稱為稚魚期。又稱變態期 (Metamorphosis)(Falk-Petersen,
2005)。在鮃鰈類,變態期非常明顯,伴隨著器官的不對性分佈並開始 底棲生活。變態期主要有兩個變化:(1). 仔魚形態特徵的消失;(2). 幼 魚特徵的出現,包括:色素樣式、體型、鰭變化 (如鯡科和鯷科)、發 光器官的形成、背棘、頭棘的退化 (鮨科、金鱗魚科)、眼睛的遷移 (鮃 鰈類)、鱗的形成 (Kendall et al., 1984)。硬骨魚在稚魚期,最大的特點 是快速的生長和發育,此時的生長主要表現為肌肉組織中蛋白的沉澱 和肌肉成分和形態的變化使此時魚體形態結構明顯改變,魚鰭條的進 一步發育完善,促使稚魚的游泳能力增強,稚魚期不僅其外部形態結 構上會發生明顯變化,個體的內部生理器官的功能趨近完善,如内分 泌系统消化系统、循環系统、免疫系统、性别决定、分化的關鍵時期。
因此稚魚階段有時亦稱為前幼魚階段 (王,2015; Hubbs, 1943; Inui et al., 1985; Miwa and Inui, 1991; Yamano et al., 1991; Miwa et al., 1992;
Tanaka et al., 1995; Padrós and Crespo, 1996; Fournier et al., 2000; Gavlik et al., 2002; Gisbert et al., 2004; Applebaum et al., 2004; De Assis et al.,
2004; Perez-Casanova et al., 2006)。
(4). 幼魚期 (Juvenile)
幼魚時期之個體和成年個體除了體型大小外,在外部形態各方面 均保持一致。幼魚期的個體性腺尚未成熟,不具備一般的副性徵,幼
14
魚期的長短和棲息場所、季節等外部環境的變化有著重要的關系。條 石鯛稚魚經常棲息於沿海水域,隨著海藻漂移,長成後遷移至岩礁地 帶 (Mundy, 2005)。Tamaki (1995) 指出條石鯛稚魚期具群遊性,在一 定水域不分散而成群生活,棲息於海藻區域時會形成較大之族群;岩 礁地區則相反,小族群會定棲於同一棲地;大族群則具遷移行為,若 成群條石鯛長時間棲息於同一地區,則會轉成數個小族群定棲。幼魚 期的個體會利用所攝取的營養物質進行蛋白儲備,使個體快速的生長 發育,由於天敵的捕食、種內的殘食、疾病發生、環境變化等外在因 素影響,幼魚階段也是死亡率高的時期 (王,2015)。
五、植物性餌料生物
微藻種類繁多,廣泛的存在於淡水、半淡鹹水及海水中,可行光 合作用且為整個生物圈中重要的初級生產者 (De Pauw and Pruder,
1984),目前普遍使用於醫學保健食品、污染防治及水產養殖中。微藻 在分類上依據所含的光合成色素、儲藏物質及細微構造可分為九大門:
藍 綠 藻 們 (Cyanophyta) 、 綠 藻 門 (Chlorophyta) 、 金 褐 藻 門
(Chrysophyta)、隱藻門 (Cryptophyta)、矽藻門 (Bacillarriophyta)、裸藻 門(Euglenophyta)、定鞭藻門 (Haptophyta)、綠鞭藻門 (Prasinophyta)、
甲藻門 (Pyrrophyta or Dinophyta)。本實驗一所使用之微藻分別為金藻 門 的 海 洋 擬 球 藻 (Nannochloropsis oculata) 與 綠 藻 門 的 周 氏 扁 藻
(Tetraselmis chui)。
1. 海洋擬球藻
海洋擬球藻(Nannochloropsis oculata)屬金藻門(Chrysophycophyta)、
針眼點藻綱 (Eustigmatophyceae)、真眼點藻目 (Eustigmatales)、單株藻 科 (Monodopsidaceae)。為小球體細胞,無細胞壁、鞭毛、眼點及葉綠 炳 (girdle lamella)構造;直徑為 2 - 5 μm。溼重檢測下其蛋白質含量占
50 %、EPA 30 %;內含色素為葉綠素 a 及類胡蘿蔔素。養殖方面多應 用於壺形輪蟲滋養及海水魚苗之投餵;亦可直接投餵當作牡蠣、文蛤、
淺蜊及真參幼生餌料。培養之鹽度範圍 20 - 35 ppt;光度為 1000 - 12000 Lux 間;溫度 10 - 35 ℃,以 25 - 31 ℃ 最佳,pH 值 7.5 - 8.8 (蘇,
1999;趙等,2002)。
2. 周氏扁藻
周氏扁藻 (Tetraselmis chui) 屬綠藻門 (Chlorophycophyta)、綠色鞭 毛藻綱 (Prasinophyceae)、綠色鞭毛藻目 (Prasinocladales)、綠色鞭毛藻 科 (Prasinocladaceae),直徑為 8 - 16 μm。藻體型態為橢圓或卵圓形,
細胞內有一杯狀並呈現綠色之色素體,其為葉綠體,細胞後端有一蛋 白核。杯狀缺口處伸出四根等長鞭毛,具備橙紅色眼點。乾重下蛋白 質可達 74 %,碳水化合物為 21 %,脂質僅含 4 %。18 : 1 n-9 含量為 33
%;EPA 含量不高,僅為 4 %。內含色素為葉綠素 a、b、β胡蘿蔔素
16
及管藻黃素 (Siphonoxanthin)。養殖應用方面常見魚投餵輪蟲及海水魚 苗;亦有投餵魚貝介類之軟體動物及種苗培育,培養溫度為 15 - 30 ℃,
以 25 ℃ 最佳;鹽度 10 - 40 ppt 下皆能增殖,30 ppt 下增殖效果最 佳。光照強度為 500 - 10000 Lux 間 (蘇,1999;趙等,2002)。
六、動物性餌料生物
水產種苗的人工繁殖過程中,由於剛孵化的魚蝦貝幼苗由於消化 系統分化不全,大多數剛孵化的魚、介、貝幼苗無法消化及代謝配方 飼料,因此種苗幼生的營養是許多魚介貝類生產工業化的瓶頸,雖在 魚苗初期階段已有開發專用配合飼料,尚無法完全取代天然餌料生物
(Watanabe and Kiron, 1994),因為天然浮游動物體內富含可消化的營養,
並含大量消化酵素,可幫助魚苗分解其攝食的食物。適當的餌料的選 擇標準為營養成分及消化酵素含量高、配合幼生之適口性、營養需求,
來源容易取得以及短時間可大量生產供應,以食用或研究中之動物性 餌料如輪蟲、豐年蝦、橈腳類、枝角類、線蟲、絲蚯蚓等,其中最普 遍應用的種類為輪蟲、橈腳類及豐年蝦 (蘇,1999)。
1. 褶皺臂尾輪蟲 (Brachionus plicatilis)
褶皺臂尾輪蟲屬袋型動物門 (Aschelminthes)、輪蟲綱 (Rataoria)、
壺型動物目 (Brachionoida)、壺型輪蟲科 (Brachionoidae) 之壺型輪蟲 屬,為一種小型動物性浮游生物,因其頭部有一轉輪狀之纖毛冠
(ciliated corona) 而得名。臂尾輪蟲 (Brachionus plicatilis) 長約 123 - 292 μm,寬約 114 - 199 μm (Snell and Carrillo, 1984),其大小適中、可 高密度培養、生產快速、對水溫及溫度耐受度高且游泳緩慢,為幼苗 不可或缺之餌料生物。趙等人 (2002) 實驗證實將其應用於鯛魚、草蝦、
斑節蝦及蟳類等水產生物均有良好之繁殖成果。影響輪蟲增殖之因素 有食物、溫度、鹽度、光、養殖環境及輪蟲生理狀況等,其中直接影 響魚苗活存率之關鍵因數為其攝食之食物。輪蟲以濾食為主,大型輪 蟲可濾食約 5 - 25 μm 之食物,極小輪蟲可濾食約 2 - 20 μm 的食物,
只要是小於 25 μm 的微藻、酵母、蛋白質為膠囊飼料、有機碎屑等皆 可被輪蟲濾食 (趙等,2002)。整體而言,餵食輪蟲的餌料大小以 15 μm 以下更為理想 (張,1997)。輪蟲的營養成分主要來自被攝入的微藻、
酵母或細菌 (Fulks and Main, 1991),然而單獨使用酵母培養的輪蟲來 投餵魚苗,會因為缺乏高度不飽和脂肪酸而有營養缺乏情形,造成魚 苗畸形或死亡率上升,此問題可利用滋養液或微藻進行二次培養做營 養強化或與微藻一起投餵來改善輪蟲品質 (Øie et al., 1994; 蘇等,
1998)。近幾年已有多位學者已發現 HUFA 對海水魚蝦苗的活存及成 長亦相當重要 (Kitajima et al., 1980; Yone, 1980; Immanuel et al., 2001 ),
特別是 ω3 高度不飽和脂肪酸 DHA (22:6ω3)、EPA (20:5ω3) 含量對 魚苗發育階段為不可缺少的重要物質(Southgate and Lou, 1995),並且其
18
無法自行合成,必須依賴外界供應,故屬於發育過程中之必需脂肪酸
(Villegas et al., 1990)。海水微藻可為輪蟲提供蛋白質、能量、多元不飽 和脂肪酸 (PUFA; polyunsaturated fatty acid)、維生素、固醇類及色素
(Tredici et al., 2009),海洋擬球藻生長快速、容易培養特性為海水魚繁 殖應用最重要的微藻,其富含 EPA,但較缺乏 DHA,因此在幼苗培 育時仍須搭配餵食其他種類微藻為佳 (Tucker, 1998)。根據輪蟲的培養 與收穫方式可分為: 批次式、連續式和半連續式三種養殖方法 (張,
1997;Snell, 1991; Fulks and Main, 1991)。
2. 短腳異劍水蚤 (Apocyclops royi)
短角異劍水蚤屬於節肢動物門 (Arthropoda)、甲殼綱 (Crustacea)、
劍 水 蚤 目 (Cyclopoida) 、 劍 水 蚤 科 (Cyclopoidae) 、 劍 水 蚤 亞 科
(Cyclopinae)、異劍水蚤屬 (Apocyclops) (鄭等,2009)。最小發育階段 之橈腳類體型約 125 μm 左右,成蟲體型約 1 - 2 mm,為海洋中野生 魚苗之重要蛋白質來源,並含豐富且完整之必需脂肪酸組成 (Tucker, 1998)。建立新物種之繁殖技術時,常使用橈腳類作為餌料 (Conceição et al., 2010)。目前台灣的橈腳類生產模式以室外養殖池為主,生產方式
有二:(1). 專業生產者:以魚漿、飼料、豆粉、雞糞等作水的方法培 養,池中混養少量的魚蝦,(2). 兼業生產者:自魚、蝦養殖池中直接 捕撈。捕撈時需在養殖池的水車前架設定置網,利用水車轉動的水流,
將池中的橈腳類帶入網內。由於橈腳類夜晚有浮游於池水中上層的習 性,一般業者大都在天亮前完成捕撈作業,隨即載送給繁殖業者在清 晨時投餵魚苗。短角異劍水蚤體型小、容易採集與培養,在良好環境 下,短時間可成熟生殖,是非常有潛力的品種 (鄭等,2009)。目前培 育海水魚苗的橈腳類多來自室外養殖池,品質與產量均不穩定,有時 還會引入其他病菌、病毒及寄生蟲等,可利用益生菌的代謝產物滋養 橈腳類,以減少感染,或以臭氧與二氧化氯充分將收穫後的橈腳類充 分清洗後,亦可以微藻充分滋養後,再行餵養,對魚苗健康與品質都 有正面助益 (鄭與陳,2009)。許多研究指出,以橈腳類幼生做為仔稚 魚餌料可提升其活存率 (陳與蘇,2005;鄭與陳,2009;江等,2012;
呂,2015)。橈腳類無節幼蟲含高度的 EPA 與 DHA,為魚類必須不飽 和脂肪酸,有利於魚苗吸收利用,因此可促進成長與提升活存率與免 疫力 (鄭與陳,2009)。
3. 豐年蝦 (Artemia)
豐年蝦屬節肢動物門 (Athropoda),鰓足亞綱 (Branchiopoda),無 甲目 (Anostraca),豐年蝦科 (Artemiidae),豐年蝦屬 (Artemia),豐年 蝦分佈於全世界高鹽度水域,對鹽度的適應範圍極廣,半淡鹹水至 90 ppt 皆可存活,目前主要產地為美國、蘇聯、中國和加拿大。市售豐年 蝦耐久卵依產地和品系的不同,其孵化率、孵化時間、無節幼蟲大小、
20
成長速率和營養組成都有所不同。一般耐久卵在水溫 28 ℃,低鹽度 0 - 20 ppt 下約 20 - 40 hr 孵化成無節幼蟲 (nauplii instar I, 430 - 515 μm),此時不會攝食。在 6 - 8 hr 後蛻變成二齡幼蟲 (instar II,
metanauplii),並開始開口進食 50 μm 以下的食物顆粒,如微藻、酵母、
細菌、原生動物和有機懸浮顆粒等 (邱,2005)。剛孵出的無節幼蟲營 養組成豐富,富含類胡蘿蔔素和高度不飽和脂肪酸,適合作為水產種 苗的營養來源 (Murugan et al., 1995;Tsushima and Matsuno, 1999)。隨 著豐年蝦品種不同,無節幼蟲營養組成也不同,其中 DHA 和 EPA 的 含量更直接影響仔魚的成長率和活存率。當蛻變成二齡幼蟲後,乾重 減少 20 %;脂肪酸減少 26 %;總能量減少 27 %,使其漸失去營養價 值。利用豐年蝦之濾食行為,使用藻類、酵母或是乳化油來強化其營 養組成 (蘇,1999;Léger et al., 1986)。
七、蛋白質之生理功能
蛋白質是魚體組織中主要的物質,約占乾重之 65 - 70 % (Wilson and Halver, 1986)。不僅是生物體各組織的組成,同時也可以代謝產生
能量,也參與許多重要生理功能,例如: 構成與維持魚體的體組織(體
組織蛋白質更新、修復及維持體蛋白現狀);生長 (生物體蛋白質的增 加);作為部分能量來源以及組成機體各種激素如賀爾蒙、酵素等具有 特殊生物學功能的物質。魚類必須不斷攝取蛋白質以獲得胺基酸,藉
以構成魚體的組織及新陳代謝,因此飼料中提供蛋白質或胺基酸是不 可或缺的 (李,1998; Jobling and wandsvik, 1983)。蛋白質基本結構是 由二十多種氨基酸結合而成,而各個胺基酸之間藉由肽鍵(peptide bond) 相互鍵結。當魚類消化蛋白質釋放游離胺基酸,以主動運輸經小腸上 皮吸收,經由門脈系統 (portal vein system) 運送到肝及其他組織,再 將這些胺基酸加以合成,作為魚體維持現狀或是提供成長所需 (Bowen,
1987)。另一方面,亦可經由體內代謝或異化作用 (catabolism) 而產生 能量 (Walton and Cowey, 1982)。蛋白質亦為飼料中最重要的營養物質 之一,而水產養殖產業所用飼料價格約佔總成本 40 - 60 %,其中又以 蛋白質來源的支出較高 (Akiyama et al., 1985),飼料中蛋白質之品質取 決於胺基酸組成與平衡,優良的蛋白質來源,需具有魚體生長所需的 氨基酸種類與含量,而不適當之蛋白質來源,造成魚類利用不佳影響 生長,這些不能消化吸收之蛋白質代謝產物排除魚養殖池中,亦造成 細菌繁衍、水質含氮廢物過高等不良後果,進而影響魚類生長 (Li and
Lovell, 1992; Lovell, 1996)。飼料中蛋白質含量過多,魚體會將蛋白質 代謝為能量用或以脂肪儲存;然而蛋白質代謝必須提供額外能量先進 行去胺作用 (deamination),排除多餘無法用於生長的氨基酸;若飼料 中蛋白質含量太少,則不足供給魚體生長所需 (Winfree and Stickney, 1981; Jauncey, 1982; Wilson, 1989)。將銀鱸 (Bidyanus bidyanus) 和何氏
22
棘鲃稚魚 (Spinibarbus hollandi) 分別餵以蛋白質含量不足之餌料,其 成長較差且魚體蛋白質含量低於正常組 (Yang et al., 2002; Yang et al.,
2003)。因此,研究飼料中蛋白質的含量對養殖生物的成長以及養殖成 本是一重要且決定養殖效益的重要因素之一。
八、魚類蛋白質的最適營養需求
魚類最適蛋白質需求量研究起源於 1958 年 De Long et al.以酪蛋 白-明膠作為蛋白來源對國王鮭 (Oncorhynchus tshawytscha) 之研究報 告。陸續有許多學者以不同蛋白質來源及含量對不同魚類進行實驗,
以決定各種魚類之最適蛋白質需求量相關研究,整理於 Table 1。魚類 因成長環境與陸生動物差異甚大,故其對各種營養物質需求亦與陸生 動物有差異。一般而言,魚類比陸上蓄養之動物需要更多蛋白質,如 家畜及家禽食物中之蛋白質含量約為 12 - 25 %,能促進成長,而魚類 對食物中蛋白質需求達 35 - 55 %,有最大成長率 (Bowen, 1987)。
魚類對蛋白質需求量受以下因素影響:
1. 魚種食性
肉 食 性 魚 類 對 蛋 白 質 的 需 求 量 比 草 食 性 或 雜 食 性 的 魚 類 高 (Yoshimatus et al., 1992),例如: 瑪拉巴石斑魚 (Epinephelus malabaricus) 及河鱸 (Perca fluviatilis) 對蛋白質的需求分別為 50.2 % 及 36.8 - 43.6 % (Fiogbe et al., 1996; Shiau and Lan, 1996),雜食性魚類如黑鯛
(Sparus macrocephalus) 及大頭鰱 (Aristichthys nobilis) 對蛋白質需求 量為 41.4 % 及 30 % (Santiago and Reyes, 1991; Zhang et al., 2010),草 食性魚類如鯉魚 (Cyprinus carpio) 的蛋白質需求量為 34.1 % (Liu et al., 2009)。另外在消化酶方面,肉食性魚類之消化蛋白酶活性比草食性
及雜食性魚類為高 (Reimer, 1982)。
2. 魚體大小
魚體成長階段不同有不同蛋白質需求量,稚魚通常將蛋白質用於 成長,所以有較高量的蛋白質需求,隨著魚體增大而減少 (Millikin, 1982; Watanabe et al., 2000),例如初重 1.5 g 的馬拉巴石斑 (Epinephelus slamonides) 之蛋白需求量為 54 % (Chen and Chen, 1986),初重 3.8 g
的馬拉巴石斑之蛋白質需求量為 47.8 % (Chen and Tsai, 1994);初重 3.1
g 之牙鮃 (P. olvaceus) 對蛋白質需求量為 50 %,初重 22.7 g 之牙鮃 對蛋白質需求量則為 45 % (Lee et al., 2002)。初重 1.56 g 及 9.49 g 之 花魚 (Hemibarbus maculates) 對蛋白質需求量分別為 37.4 % 及 35.5
% (Chen et al., 2010),初重 114 g 與 14 g 之鉗魚 (Ictalurus punctatus) 蛋白質需求量分別為 25 % 及 35 % (Page and Andrews, 1973)。
3. 生長環境
魚類蛋白質需求會受水溫變化的影響,大鱗鮭魚 (chinook salmon) 在 8 ℃ 及 14 ℃ 下之最適蛋白質需求分別為 40 % 及 55 % (De
24
Long et al., 1958) ; Millikin (1982; 1983) 指 出 條 紋 狼 鱸 (Morone saxatilis) 於 20 ℃ 下蛋白質需求為 47 %;24 ℃ 下蛋白質需求量為
55 %。亦有學者探討鹽度對魚類最適蛋白質需求量影響;Zeitoun et al.
(1973) 指出虹鱒 (Salmo gairdneri) 之最適蛋白質需求量,鹽度 10 ppt 最適蛋白質需求量為 40 %,鹽度 20 ppt 最適蛋白質需求量為 45 %;
然而餵食吳郭魚 (Tilapia, O. niloticus × O. aureus) 24 % 蛋白質飼料時,
無論是淡水 (Shiau et al., 1987) 或海水 (Shiau and Huang, 1989) 試驗,
其增重率皆無差別。
4. 飼料組成
飼料中能量含量會影響魚類對蛋白質之攝取 (Kaushik and Médale, 1994),當飼料能量不足時,魚體會代謝蛋白質作為能量使用,使蛋白 質無法完全利用於成長或維持;飼料中能量過高時,會造成魚類容易 產生飽食感而減少攝食,造成攝取的蛋白質無法達到最佳成長,可藉 由 添 加 碳 水 化 合 物 或 脂 質 來 達 到 蛋 白 質 節 約 效 應 (protein-sparing effect) (Hernandez et al., 1995)。
九、魚粉概況
水產飼料為養殖主要生產成本 (Pandian, 1989; Cheng et al., 2003),
其中魚粉為人工飼料中蛋白質主要原料,亦是最昂貴的成分 (Akiyama et al., 1991; Lim, 1996; Lemos et al., 2000; Martinez et al., 2002),也是成
長影響最大因素 (Andrew et al., 1972; Balaz, 1973; Alava and Lim, 1983;
Kureshy and Davis, 2002)。全世界主要魚粉生產國與輸出國分別為祕魯、
智利、丹麥、冰島、挪威、南非、美國及日本。畜產及養殖產業的飼 料費用佔其生產成本比例非常高,分別佔約 40 % 及 65 % (江和陳,
2000)。整隻為切割之全魚或切割後之魚塊、魚雜,新鮮為分解者均可 當魚粉原料,乾燥之即為魚粉。傳統以直火法製造,對魚粉品質影響 大;今多利用蒸氣間接加熱法或是低溫真空乾燥法以保存魚粉營養。(林,
2008)。魚粉的適口性佳、營養豐富、消化率高、胺基酸組成平衡,並 具 有 B2 、 B12 等 維 生 素 及 未 知 生 長 因 數 (UFG, unknow growth factor)(洪,1980),然而,根據聯合國農糧組織 (Food and Agriculture Organization of the United Nations, FAO) 1961 - 2006 年的統計,魚粉產 量維持魚每年 500 萬至 700 萬噸之間,直至 2008 年世界五大主要 出口國 (祕魯、智利、美國、挪威、冰島) 的魚粉產量均有下降,比 2007 年減產約 100 萬噸,未來也無法使魚粉產量上升 (謝,2010)。國際魚 粉及魚油組織 (International Fish meal and fish oil organization, IFFO) 指出 2014 年魚粉產量為 414 萬噸,其中 35 % 為魚類加工後的副產 品。且比例持續上升,過去主要魚粉生產國祕魯的魚粉價格為每公噸 1200 美元,由於 2012 年祕魯鯷魚魚貨產量減少,導致魚粉在 2013 年 1 月達到每公噸 2150 美元 (國際漁產資訊 246 期,2013)。從 2013 年
26
1 月到 2014 年 1 月價格約減少 20 %,不過價格仍維持在高檔 (國際 漁產資訊 264 期,2014),養殖漁業產量的持續增使飼料魚粉的需求增 加,因此針對不同魚種營養需求了解最適生長飼料蛋白質含量以提升 養殖效益。
材料方法
一、實驗設計
本論文分為二個部分: 實驗一、斑石鯛 (Oplegnathus puctatus)人工 孵化繁殖技術建立;實驗二、不同蛋白質含量對斑石鯛幼魚成長之探 討。
實驗一、斑石鯛 (Oplegnathus puctatus)人工孵化繁殖
1. 種魚培育與受精卵採集
本次孵化試驗種魚取自高雄市林園區合作養殖戶,使用以鱗片紋 路鑑定 6 年齡斑石鯛種魚,檢疫 3 天後將種魚各自蓄養於 8 噸圓形 玻璃纖維養殖槽,共 2 池,水溫維持在 20 ± 1 ℃,鹽度範圍在 33 - 35
ppt,養殖缸上方設置蘭花網遮蓋池面約 90 % 面積。每週清潔養殖缸 一次並於每日清晨抽底清除缸底髒物、更換海水以維持缸內水質穩定,
每日以胭脂蝦 (Aristaeomorpha foliacea)、鯖魚 (Scomber scombrus)、
南魷 (Sthenoteuthis oualaniensis) 及生鮮牡蠣等混合投餵 1 次。生殖季 期間發現斑石鯛具備婚姻色時,以手指輕壓種魚腹部檢查是否有精液 流出作為雄魚判斷依據;雌魚為腹部飽滿個體及以塑膠軟管插入生殖 孔吸取是否有卵粒作為判斷依據。雄魚為體長 25 - 40 cm,體中 2.5 - 3.5 kg 之自然成熟個體;雌魚挑選體長 25 - 40 cm,體重 2.5 - 3.5 kg 之個 體,先以塑膠軟管插入生殖孔吸取卵粒後觀察其成熟度,再以人類絨
28
毛膜促性腺激素 (HCG) 1000 I.U. / kg、促黃體素釋放素 (LHRH - A2) 20 μg / kg 分別以生理食鹽水溶解後於背部肌肉採一次性注射催熟,
24 hr 後再注射一次上述二種激素觀察腹部是否肥滿、生殖孔是否突出 之個體作為採集成熟卵之依據。生殖季節傍晚晴朗時期將斑石鯛雌魚 與雄魚麻醉後分別取卵與精子,將精子以少量海水稀釋後與卵混合,
以毛筆緩慢攪動靜置 1 min,重複上述步驟 3 次後將卵以浮游生物網 收集,以海水洗去多餘精子後在將上浮於中、表層受精卵置入 100 L 容 器打氣,隔日清晨將受精與未受精卵分別收集秤重以瞭解斑石鯛人工 受精率。
2. 餌料生物與人工飼料
實驗用之海洋擬球藻、周氏扁藻種原購自國立台灣海洋大學水生 動物實驗中心,培養採半連續式培養 (Semi-continue culture),經濾袋 過濾後將海水按比例加入 Walne 培養液配方 (Walne, 1974),置入 20
L 透明酒果筒以高溫高壓滅菌,待冷卻後再依比例添加藻種後打氣培 養。光照週期維持 L 16 : D 8,照度 3000 - 5000 Lux,溫度維持 27 ± 0.5
℃,經 3 - 5 天呈現濃綠色時即可採收使用或繼續擴陪。
輪蟲體型約 130 - 300 μm,以 10 噸長方形玻璃纖維槽培養,加 入經濾袋過濾海水後,以淡水將鹽度調至 25 ppt,水溫維持 27 ± 0.5 ℃,
添加發酵液後 3 - 4 天後水色轉紅褐色後接種輪蟲,以微打氣方式培養,
每日添加適量發酵液維持水色,一週後即可以 300 目浮游生物網收集 輪蟲,依幼苗不同口徑大小使用不同網目 (150、120、100、80 網目) 篩選。
橈腳類培養方式參照輪蟲之培養方法,橈腳類需要較高溶氧,因 此培育期間採強打氣方式養殖,溶氧維持 > 5 ppm,一週後即可以 300 目浮游生物網收集橈腳類,依幼苗不同口徑大小使用不同網目 (150、
120、100、80 網目) 篩選橈腳類幼蟲及成蟲。
豐年蝦耐久卵為 Sanders 牌之 Great Salt Lake 品種 (Artemia franciscana),去殼流程如下: 將卵浸泡於微打氣之淡水 1 hr,靜置沉
澱後,將上方懸浮卵倒除後收集下沉卵,以每克的耐久卵加入 13 ml 的漂白水溶液 (淡水/漂白水 = 1/1 比例配置) 作為去殼溶液,並攪拌觀 察耐久卵顏色變化,顏色呈現橘色時,使用浮游生物網收集並以乾淨 自來水清洗數次後再添加硫代硫酸鈉去除殘留氯,最後將卵風乾後存 於 4 ℃ 冰箱備用,孵化時將去殼卵放入強烈打氣之海水中,水溫 27
℃ 下約 16 - 18 hr 即可孵出無節幼蟲,孵化後即可採收並投餵幼苗,
多餘豐年蝦存於 4 ℃ 冰箱備用並於 12 hr 內用畢。
幼苗培育期間投餵之黑粒粉、海藻粉購自群冠公司。
3. 受精卵與胚胎發育
實驗期間共人工採卵 2 次,採集受精卵後置入 300 L 玻璃纖維桶
30
中以連續微打氣、止水狀態做為孵化環境,孵化期間取出部分受精卵 置入 500 ml 燒杯中,燒杯外隔水加溫使溫度維持於 23 ± 1 ℃,每隔
30 min 利用塑膠滴管自燒杯內吸取受精卵至凹槽玻片上並以水蓋過受 精卵後,以複式顯微鏡觀察胚胎發育,並以數位相機拍照後再經影像 分析軟體 (Image J) 分析並記錄受精卵之卵徑、油球徑與胚胎發育各期 之時間與型態特徵。胚胎發育時期經歷時間,以顯微鏡視野中有 50 % 的受精卵胚胎從前一階段至下一階段所經歷的時間做判讀依據。
4. 仔稚魚孵化繁殖與型態觀察
採集受精卵後,隔日清晨將受精卵移入 4000 L、水溫 26 ± 1 ℃、
鹽度 35 ppt,pH 8.0 - 8.2 之方形玻璃纖維槽做為幼苗培育環境。仔魚 餌料以經海洋擬球藻及周氏扁藻滋養 3 hr 之輪蟲、橈腳類、豐年蝦及 人工配合微粒飼料做投餵餌料,每日視水色濃度適當添加藻水,並輔 以 250 目浮游生物網袋搓洗後之海藻粉及黑粒粉,將其均勻潑灑至養 殖槽中維持水色。育苗初期每日加入光合菌 (Rhodobacter capsulatus)、
枯草桿菌 (Bacillus substilis)、希瓦氏菌 (Shewanella sp.) 各 2 ppm 維 持水質穩定 (氨-氮 < 0.25 ppm;亞硝酸-氮 < 20 ppm);育苗中後期開 始少量流水養殖,一週後將幼苗移至新養殖槽蓄養。培育期間定期採 樣 5 尾斑石鯛幼苗,隨魚體大小不同而分別以複式顯微鏡及解剖顯微 鏡觀察及拍攝記錄魚苗之體色及形態特徵變化,拍攝前以麻醉劑處理
幼苗後,待其跳動趨緩再移至顯微鏡進行拍照記錄,並以電子游標尺 或尺規測量平均體標準長,了解斑石鯛幼苗生長情形。
二、實驗二 不同蛋白質含量對斑石鯛成長之探討
實驗二探討以不同蛋白質含量的飼料搭配等油脂等能量的飼料,
投餵初重約 16 g 實驗一人工繁殖的斑石鯛 (Oplegnathus punctatus) 探討其成長表現與體組成影響。
1. 養殖生物
斑石鯛幼魚為人工孵化繁殖 8 週,將健康個體挑選後安置於 4000 L FRP 桶中,並以台榮公司市售鰻魚飼料 (粗蛋白含量 45 %) 餵 食馴養 2 週,待幼魚適應養殖環境與接受自製人工配合飼料後分別秤 重,挑選平均體重約 16 g 個體進行實驗。
2. 養殖條件
實驗系統為 18 個 120 L 玻璃水族箱組成的流加水養殖系統,引 入的海水設置濾袋做初步物理過濾,每缸水溫維持於 29 ± 1 ℃,鹽度
34 - 35 ppt,pH 8.1 - 8.3,光週期採自然光照,每日早上餵食前虹吸底 部沉積物,並且換除 1/2 海水維持穩定水質,每 1 週清潔一次缸壁維 持養殖環境穩定。光週期採自然光照,每日於早晨 9 : 30 及下午 6 : 00 分別投餵 1 餐至飽食,並且記錄每組攝食量。每 2 週記錄一次體全 長及體重;實驗為期 8 週,第 8 週實驗結束後採樣進行魚體分析。
32
3. 飼料配製
實驗二飼料配方見 Table 2,每種原料秤重裝袋,再依克數由小至 大均勻混合後過篩,實驗飼料以紅魚粉 (brown fish meal) 為主要蛋白 源,搭配 7 % 鱈魚肝油/玉米油 (2:1) 為油脂來源,並以 α- starch 為 黏著劑,再以 α- cellulose 調製成等能量之飼料,配置蛋白質含量為
35 %、40 %、45 %、50 %、55 %及 60 % 六組實驗飼料,分別由 A1 - A6 標示,除了鱈魚肝油與玉米油,其它原料皆以 60 網目篩網過篩並均勻 混合後,在加入鱈魚魚油與玉米油及約原料重量的 35 - 40 % 蒸餾水 均勻混合並攪拌成團狀,以絞肉機擠成條狀後經烘箱 57 ℃ 烘乾 24
hr 後,以粉碎機粉碎成 4.8 mm 大小顆粒狀,製粒完成後保存於 - 20
℃ 冰箱內,投餵前取出放置室溫 20 min 後再行投餵。
4. 分析項目
實驗飼料及魚體組成成分分析,包含水分、灰分、粗蛋白、粗脂 質等,均參照 A.O.A.C (1995) 之方法分析,魚體組織樣品待實驗結束 後先禁食 24 hr,再以碎冰凍暈,並個別稱重及片取肌肉組織。肌肉組 織的取樣方法為頭部至尾部二側的肌肉。取下的肌肉去除魚皮,以均 質機均勻絞碎後以 105 ℃ 烘乾,烘乾後的樣品組織再以均質機均勻 絞碎後,冰存於 - 20 ℃ 冰箱,待分析時再取出。
(1). 水分測定 (Moisture)
取經磨碎後的樣品 2 g 放入已秤重且已達恆重的乾燥坩鍋中,再將其 置於 105 ℃ 恆溫乾燥箱,每 1 hr 取出放在乾燥器中冷卻至室溫後秤 重,重量記錄至小數點後 3 位,直至恆重為止。
計算公式:
Moisture (%) = (W1 - W2) / (W1 - W0) × 100 % W0: 坩鍋的恆重 (g)
W1: 坩鍋重+未烘乾樣品重量 (g) W2: 坩鍋重+烘乾後樣品重量 (g)
(2). 灰分測定 (Ash)
取經磨碎後的樣品 1 g 至小數點後 4 位,置入已秤重且達恆重的乾燥 坩鍋中,在將坩鍋加蓋放入灰化爐中,以 550 ℃ 灰化 15 hr,將樣品 灰化製成灰色粉末狀為止,待溫度降至 100 ℃ 後,將坩鍋取出置於 真 空 乾 燥 器 中 , 待 溫 度 降 至 室 溫 秤 其 重 量 , 重 量 紀 錄 至 小 數 點 後 3 位。
計算公式:
Ash (%) = (W1 - W0) / S × 100 % W0: 坩鍋的恆重(g)
W1: 坩鍋重+灰化後樣品重量 (g) S: 樣品重量 (g)
(3).粗蛋白測定 (Crude Protein)
34
粗蛋白依照 Micro-Kjeldahl (A.O.A.C., 1995) 方法測定,秤取樣品 0.5 g 至小數點後 4 位,將其置入凱氏氮管,並加入催化劑 5 g (K2SO4:
CuSO4‧5H2O = 9:1),隨後加入 15 ml 濃硫酸,將凱氏氮分解瓶放入 粗蛋白消化爐 (Automatic Kjeldahl Digestion Units - DKL Series, Velp
Scientifica) 加熱至 380 ℃ 使樣品分解至淡綠色澄清狀,取出冷卻至 室溫後,加入 70 ml 蒸餾水及 15 ml 濃度為 35 % NaOH 水溶液,再 以全氮蒸餾器 (UDK 129 Kjeldahl Distillation Unit, Velp Scientifica) 使 其加熱蒸餾出氮氣,以 25 ml 濃度 4 % 硼酸收集蒸餾液至 150 ml,
以 Methyl red 及 Bromocresol green 做為指示劑,並以 0.1 N HCl 滴定 至淡粉紅色作為滴定終點。
計算公式:
Crude Protein (%) =【(b - a) × 0.1 × 14.007 × 6.25】/ S × 100 % a: 空白組滴定量 (ml)
b: 樣品滴定量 (ml) S: 樣品重量 (g)
(4). 粗油脂測定(Crude Lipid)
使用脂肪抽出器 (Tecator Soxtec System HT 1043 extraction unit)測定,
測定前將鋁杯洗淨並置於烘箱以 105 ℃ 乾燥 1 hr,取出後至於恆溫 乾燥箱中 30 min 後秤重,並在鋁杯中加入 45 ml 的乙醚,秤取 0.5 g
樣品置於棉筒中上層並鋪上脫脂棉,設定機器溫度為 90 ℃,將棉筒 放入裝有乙醚的鋁杯中,浸泡 15 min,再利用滴濾方式滴濾 1 hr,關 起滴濾裝置,再開啟抽風空氣 15 min,使鋁杯中的乙醚揮發,回收乙 醚,取出鋁杯至於烘箱以 105 ℃ 乾燥 1 hr,取出後至於恆溫乾燥箱 中 30 min 後秤重,秤重後減去重量差即得粗脂質含量。
計算公式:
(W2 - W1) / S × 100 % W2: 恆重後脂鋁杯重 (g) W1: 萃取恆重後之鋁杯重 (g) S: 樣品重量 (g)
5. 成長指標分析
(1). 增重百分率 (Percent Weight Gain)
實驗結束後,斑石鯛幼魚的體重增加量,除以初重量,再乘以 100。
計算公式:
Weight gain (%) = (Final weight - Initial body weight) / (Initial body weight) × 100 %
(2). 飼料轉換率 (Feed Conversion Ratio, FCR)
實驗期間,斑石鯛幼魚每增加 1g 體重,所攝食的飼料乾物重。
計算公式:
Feed conversion ratio (FCR) = Feed intake / (Final body weight - initial
36
(3). 特殊成長率 (Specific Growth Rate, SGR)
Specific Growth Rate (SGR) =【ln (Final body weight) – ln (Initial body weight)】× 100 % / day (s)
(4). 攝食量 (Feed Intake, FI)
實驗期間斑石鯛攝入的飼料量。
(5). 蛋白質效率比 (Protein Efficiency Ratio, PER)
Protein Efficiency Ratio (PER) = (Final weight - Initial weight) / Feed intake × protein percent
(6) 活存率(Survival Rate)
實驗結束時,斑石鯛存活之尾數佔實驗開始時之尾數的百分比。
計算公式:
Survival rate (%) = (Final number of fish) / (Initial number of fish) × 100
%
6. 體測量指標
(1). 肥滿度(Condition Factor, C.F.)
計算公式:
Condition Factor, C.F. = Body weight / (Body length)3 × 100 %
(2). 內臟體比 (Viscerasomatic Index, VSI)
計算公式:
Viscerasomatic index, VSI = Visceral weight / Body weight × 100 %
(3). 肝體比 (Hepatosomatic Index, HSI)
計算公式:
HSI (%) = Liver weight / Final body weight × 100 %
7. 統計分析
實驗數據以 SAS / PC (SAS / PC version, SAS Institute, Cary, CA, USA)軟體進行單變異數分析(Analysis of variance, ANOVA, one-way) 測試個實驗組是否有差異,若有差異再以 Duncan’s new multiple range test 比較各處理組間之差異顯著性,顯著水準定為 p < 0.05。
38
結果
實驗一、斑石鯛胚胎發育與仔稚魚幼苗培育
1. 種魚培育與人工授精
本次實驗結果發現斑石鯛雄魚生殖季時體色及吻端呈現黑色或灰 白色;雌魚吻端亦為黑色或灰白色,且腹部稍有膨大情形 (Fig. 1),生 殖季期間將雌斑石鯛取卵觀察卵粒發現已有卵黃堆積 (Fig. 2),再以人 類絨毛膜促性腺激素 (HCG) 1000 I.U./ kg、促黃體素釋放素 (LHRH -
A2) 20 μg/ kg 分別用生理食鹽水溶解後於背部肌肉注射催熟,隔日凌 晨觀察其腹部明顯膨大 (Fig. 3),當日晚間觀察其生殖孔有明顯紅腫突 出 (Fig. 4),輕壓即可採集大量成熟卵粒,以離心管採集精子後將其與 卵以半乾式人工授精均勻混合攪拌 5 分鐘後再以乾淨海水洗去多餘 精子及完成受精過程。自 4 月初至 5 月初共進行 2 次人工授精實驗,
平均受精率為 71.20 ± 0.10 %。
2. 受精卵與胚胎發育
胚胎發育時間與特徵見 Table 3 與 Figure 5 - 7;詳細敘述如下:
受精卵為單一油球,屬於圓球狀、分離之浮性透明卵,淡黃色並富彈 性,平均直徑為 1.03 ± 0.04 mm;油球徑為 0.23 ± 0.01 mm。水溫 23 ± 1
℃ 下,受精後 40 min,受精膜吸水膨脹,動物極形成胚盤 (Blastoderm),
側面觀可見胚盤如高帽狀隆起 (Fig. 5a);受精後 1 hr 10 min 進入 2
細胞期,形成 2 個對等大小的細胞 (Blastomeres) (Fig. 5b);受精後 1
hr 25 min 與第一次分裂溝垂直的位置形成第 2 次分裂,進入 4 細胞 期 (Fig. 5c);受精後第 1 hr 50 min 形成第 3 次分裂,進入 8 細胞期
(Fig. 5d),受精後 2 hr 15 min 第 4 次分裂,進入 16 細胞期,2 個分 裂面都與第 1 次平行,形成 2 排 8 個細胞 (Fig. 5e);受精後 2 hr 45
min 發生第 5 次分裂,進入 32 細胞期,卵裂細胞大小不等,排列呈 不規則 (Fig. 5f);受精後 3 hr 25 min 進入 64 細胞期,發生第 6 次 卵裂,此時細胞分裂不完全同步,大小不等且排列不規則 (Fig. 5g);
受精後 3 hr 40 min 進入桑椹期 (Morula stage),細胞繼續在動物極快 速分裂並開始堆疊,細胞數量難以計數且分裂面難以分辨,整個細胞 團近似圓形,從側面觀察與桑椹相似 (Fig. 5h);受精後 4 hr 10 min 進 入高囊胚期 (High blastula stage),胚層形成,在動物極堆積成高帽狀囊 胚 (Fig. 5i),受精後 4 hr 45 min 進入低囊胚期,隆起細胞部分變低並 向四周擴展,囊胚層邊緣細胞 (Marginal cells) 與卵黃細胞融合形成卵 黃多核層 (Syncytial layer),胚盤開始向植物極下包 (Fig. 6a);受精後 7
hr 50 min 進入原腸早期 (Early gastrula stage),部分細胞內卷形成一個 環狀的胚環 (Germ ring) (Fig. 6b),此時胚層下包至卵黃 1/3 (Fig. 6c);
受精後 10 hr 20 min 進入原腸中期 (Middle gastrula stage),胚環更明 顯,細胞繼續下包和內卷,形成胚盾 (embryonic shield) (Fig. 6d),胚層