國立臺灣大學生物資源暨農學院昆蟲學研究所 碩士論文
Graduate Institute of Entomology College of Bioresources and Agriculture
National Taiwan University Master Thesis
南黃薊馬 (纓翅目:薊馬科) 媒介西瓜銀斑病毒之傳播模式及病 毒對薊馬生活史特徵之影響
Transmission Mode of Watermelon silver mottle virus (WSMoV) by Thrips palmi Karny (Thysanoptera: Thripidae) and Effects of
WSMoV on Life History of Thrips 陳惟德
Wei-Te Chen
指導教授:蔡志偉 博士 Advisor: Chi-Wei Tsai, Ph.D.
中華民國 101 年 7 月
July, 2012
i 口試委員審定書
ii 誌謝
本研究的完成,首要感謝我的家人經濟上的支持,讓我得以衣食無虞的 完成我的碩士論文。再來感謝我的指導老師蔡志偉老師,極有耐心指導我,
以及提供所有我需要的資源,讓我有極大的進步,且了解自己的極限。並感 謝病媒昆蟲學研究室現在及過去全體成員對我的支持。另外感謝黃莉欣博士 在論文 口試時 提出 諸多相 當中 肯及實 用的 建議 以 及在我 進行 實驗時 提供部 分材料,洪挺軒老師提醒了許多病理方面的專業知識令我受益良多。柯俊成 老師提 醒了許 多關 於中文 寫作 的注意 要點 與我遺 漏掉的 南黃 薊馬的 部分 基 本背景知識,最後個別感謝許洞慶老師、吳文哲老師、柯俊成老師、張慧羽 老師、張俊哲老師、許如君老師、奧山利規老師在我的碩士時光給予我的所 有直接的支持或間接的幫助。
以上
學生陳惟德謹誌
iii 中文 摘要
南黃薊馬 (Thrips palmi) 除了為葫蘆科作物重要害蟲外,亦傳播多種番 茄 斑 萎 病 毒 屬 (Tospovirus) 病 毒 , 例 如 西 瓜 銀 斑 病 毒 (Watermelon silver mottle virus, WSMoV) 。本研究確認南黃薊馬傳播 WSMoV 感染西瓜的傳 播機制,研究結果指出南黃薊馬可於一齡幼蟲期、二齡幼蟲期 及成蟲期獲得 病毒及傳播病毒,但主要還是於一齡幼蟲期獲得病毒,待發育至成蟲期時傳 播病毒。使用反轉錄 聚合酶連鎖反應 (reverse transcription-polymerase chain reaction) 偵 測 到 表 現 WSMoV 非 結 構 蛋 白 (nonstructural protein) 之病毒 互補 RNA (viral complementary RNA) 之 RT-PCR 產物,確認 WSMoV 在 南 黃 薊 馬 體 內 進 行 複 製 。 綜 合 以 上 結 果 證 實 南 黃 薊 馬 以 持 續 性 增 殖 型 (persistent-propagative) 傳 播模 式傳播 WSMoV。病 毒與薊馬 的 交 互 作 用 方 面,健康薊馬與感染 WSMoV 的薊馬之幼蟲存活率及發育速率 沒有顯著差 異,顯示 WSMoV 對南黃薊馬 幼蟲存活率及發育速率沒有直接影響 。當感 染 WSMoV 薊馬幼蟲取食受 WSMoV 感染的西瓜葉時,其幼蟲存活率與取 食 健 康 西 瓜 者 相 比 並 無 差 別 , 但 發 育 速 率 較 於 取 食 健 康 西 瓜 者 高 , 顯 示 WSMoV 會透過寄主植物影響南黃薊馬幼 蟲之發育速率。在不同性別 薊馬成 蟲的取食偏好性實驗中,雌、雄薊馬皆偏好取食遭薊馬取食 的西瓜植株,但 僅雄薊馬偏好 薊馬接種感染 WSMoV 之西瓜植株。這樣的結果顯示,遭薊 馬取食 及 WSMoV 的感染可增加植株對南黃薊馬 的吸引力。本研究闡述南 黃薊馬 傳播 WSMoV 感染西瓜 的傳播機制及病毒與薊馬 間交互 作用,但有 關 WSMoV 與南黃薊馬之間交互作用的細節,仍須進一步研究探討。
關鍵字:番茄斑萎病毒屬病毒、持續性增殖型、存活率、發育速率、取食偏 好性
iv 英文摘要
The melon thrips (Thrips palmi) is a pest on cucurbit crops but also a vector that transmits tospoviruses, for example Watermelon silver mottle virus (WSMoV).
In this study, the transmission mode of WSMoV by T. palmi was clarified. The results of transmission assay indicate that first -instar larvae, second-instar larvae and adults of T. palmi acquired the virus and transmitted the virus. However, adults had the highest transmission efficiency when they acquired the virus at first -instar stage. Using reverse transcription-polymerase chain reaction, the WSMoV M RNA’s complementary RNA of which ecodes virus nostructural protein was detected in virus-infected thrips. These results suggest that T. palmi transmits WSMoV in a persistent-propagative manner. Further, the effect of WSMoV infection on survival rate and developmental time of T. pami larvae was examined.
The developmental time and survival rate of WSMoV-infected T. palmi larvae were not different from those of healthy thrips. When T. palmi fed on the WSMoV-infected plants, either healthy thrips or virus -infected thrips grew faster than thrips fed on healthy plants. These results imply that WSMoV did not directly affect T. palmi, but WSMoV infection affects the developmental time indirectly through the WSMoV-infected plant. In feeding preference test of T. palmi adults, both male and female thrips prefered thrips-damaged plant and more males stayed on WSMoV-infected watermelon seedling, but females did not. Theese results indicate T. palmi is attracted by thrips damage and WSMoV infection. In this study, I not only exanined the transmission mode of WSMoV by Thrips palmi, but also studied on the interaction between virus and thrips vector. H owever, the effect of WSMoV infection on T. palmi should be studied in details to clarify the actual relationships between them.
v
Key words: Tospovirus, persistent-propagative, survival rate, development al time, feeding preference
vi 目錄
口試委員審定書 ... i
誌謝 ... ii
中文摘要 ... iii
英文摘要 ... iv
目錄 ... vi
圖目錄 ... viii
表目錄 ... ix
前言 ... 1
材料與方法 ... 5
1. 南黃薊馬、植物及病毒 ... 5
2. 西 瓜 銀 斑 病 毒 的 檢 測 ... 7
3. 南黃薊馬傳播西瓜銀斑病毒的傳播模 式 ... 8
3.1 非持續性傳播模式 ... 9
3.2 半持續性傳播模式 ...10
3.3 持續性傳播模式 ... 11
4. 西瓜銀斑病毒於南黃薊馬體內的複製 ...12
5. 西瓜銀斑病毒對南黃薊馬幼蟲發育速 率 及存活率影響 ...13
5.1 直接效應 ...13
5.2 間接效應 ...13
6. 南黃薊馬成蟲之取食偏好性 ...14
7. 統計分析 ...14
結果 ...15
1. 南黃薊馬傳播西瓜銀斑病毒的傳播模 式 ...15
1.1 非持續性傳播模式 ...15
vii
1.2 半持續性傳播模式 ...15
1.3 持續性傳播模式 ...15
2. 西瓜銀斑病毒於南黃薊馬體內的複製 ...17
3. 西瓜銀斑病毒對南黃薊馬幼蟲發育速 率 及存活率之影響 ...18
3.1 直接效應 ...18
3.2 間接效應 ...19
4. 南黃薊馬成蟲之取食偏好性 ...24
討論 ...28
1. 番茄斑萎病毒屬病毒的 傳播模式 ...28
2. 番茄斑萎病毒屬病毒對薊馬存活率與 發 育速率之影響 ...29
3. 南黃薊馬成蟲之取食偏好性 ...31
4. 結論 ...33
參考文獻 ...34
viii 圖目 錄
圖一、圓葉煙草 (Nicotiana benthamiana) ... 6 圖二、西瓜苗 (Citrullus lanatus) ... 7 圖三、西瓜銀斑病毒 (Watermelon silver mottle virus, WSMoV),於西瓜及南
黃薊馬 (Thrips palmi) 體內複製 ...17 圖四、幼蟲存活率 (Kaplan–Meier) ...19 圖五、幼蟲存活率 (Kaplan–Meier) ...21 圖六、南黃薊馬 (Thrips palmi) 幼蟲飼育於西瓜葉至前蛹期的平均發育時間 (平均 ± 標準差) ...23 圖七、南黃薊馬 (Thrips palmi) 成蟲對薊馬取食西瓜及健康西瓜植株的取食
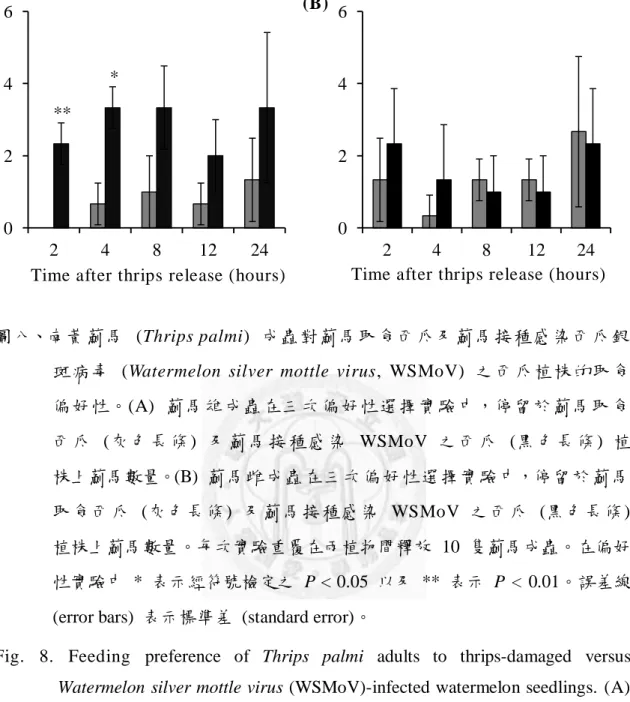
偏好性 ...25 圖八、南黃薊馬 (Thrips palmi) 成蟲對薊馬取食西瓜及薊馬接種感染西瓜銀
斑病毒 (Watermelon silver mottle virus, WSMoV) 之西瓜植株的取食偏 好性 ...26 圖 九 、 南 黃 薊 馬 (Thrips palmi) 成 蟲 對 薊 馬 接 種 感 染 西 瓜 銀 斑 病 毒 (Watermelon silver mottle virus, WSMoV) 之西瓜及健康西瓜植株 的取食 偏好性 ...27
ix 表目 錄
表一、南黃薊馬 (Thrips palmi) 傳播西瓜銀斑病毒 (Watermelon silver mottle virus, WSMoV) 傳播效率 ...16 表二、西瓜銀斑病毒 (Watermelon silver mottle virus) 的感染對南黃薊馬
(Thrips palmi) 於花豆苗上的幼蟲存活率與 幼蟲發育至前蛹期時 間的直 接效應 ...18 表三、西瓜銀斑病毒 (Watermelon silver mottle virus, WSMoV) 對南黃薊馬
(Thrips palmi) 於西瓜苗上的幼蟲發育至前 蛹期時間的間接效應 ...22
1 前言
臺 灣 氣 候 溫 暖 適 合 瓜 類 作 物 生 長 , 其 中 最 為 重 要 的 莫 過 於 西 瓜 (Citrullu s lanatu s ) 以及洋香瓜 (Cucumis melo) (Peng et al., 2011)。但在瓜 類作物的生產過程中,蟲媒植物病毒的感染,常造成產量的嚴重損失,例如:
2006 年臺灣台南地區的洋香瓜因受南黃薊 馬 (Thrips palmi) 所傳播的西瓜 銀斑病毒 (Watermelon silver mottle virus, WSMoV) 感染,造成高達八百萬 美元損失 (鄧,2011; Peng et al., 2011)。在田間,病媒昆蟲以各種不同的傳 播模式 (transmission mode) 傳播植物病毒。病媒昆蟲傳播病毒的傳播模式,
依 據 病 媒 昆 蟲 的 獲 毒 停 留 時 間 (acquisition access period, AAP) 、 潛 伏 期 (latent period) 與可持續傳播植物病毒的時 間 (persistent period),概分成以 下三 類: (1) 非持續性傳播 (nonpersistent transmission);(2) 半持續性傳播 (semipersistent transmission);(3) 持續性傳播 (persistent transmission) (Nault, 1997)。 持續性 傳 播模 式 又 可 依病 毒 是 否 於病 媒 體 內 複 製 、增殖 , 細分為持 續性循環型 (persistent-circulative) 與持續性增殖型 (persistent-propagative) (Hougenhout et al., 2008) 。 大 多 數 的 蟲 媒 植 物 病 毒 乃 依 靠 半 翅 目 昆 蟲 (Hemiptera) 或 纓 翅 目 昆 蟲 (Thysanoptera) 傳 播 (Hougenhout et al., 2008)。
薊馬屬纓翅目昆蟲,是目前唯一 以持續性增殖型傳播模式傳播布尼亞病 毒 科 (Buyaviridae) 的 番 茄 斑 萎 病 毒 屬 (Tospovirus) 病 毒 的 病 媒 昆 蟲 (Whitfield et al., 2005)。但是薊馬與其他以 持續性傳播模式傳播 病毒的病媒 昆蟲不同之處在於,薊馬需要於幼蟲期獲得病毒才有機會於成蟲期傳播病毒 (Ulman et al., 1992; van de Wetering et al., 1996)。當薊馬幼蟲取食染 病植物 的 汁 液 時 , 會 一 併 攝 入 番 茄 斑 萎 病 毒 顆 粒 時 , 病 毒 顆 粒 則 會 經 由 前 腸 (foregut) 進 入中 腸 (midgut), 與中腸 細 胞 上 的 病毒 受體進行結 合 , 然 後感 染中腸腸道微絨毛 (microvillae) 於其細胞內增殖,離開中腸腸道微絨毛細 胞後再進入環繞腸道之肌肉細胞,爾後進入血淋巴 (hemolymph) 接著感染 其它組織,最後進入唾液腺 (salivary gland),進而在薊馬唾液腺內進行複製 造成唾腺感染 (Ulman et al., 1993)。另有學者指出番茄斑萎病毒在薊馬體內
2
的感染,主要是經由感染連接中腸與唾液 腺的韌帶 (ligament),造成唾液腺 感染,這種路徑遠比經由血淋巴來得快速且 有效率 (Nagata et al., 1999)。在 病毒感染薊馬唾液腺後,當薊馬成蟲取食健康植株時,注入植物細胞的唾液 含有病毒顆粒,接種病毒到健康植物 (Whitfield et al., 2005)。
蟲媒植物病毒要成功 地傳播,除了病媒昆蟲本身與病原兩者的交互作用 外,病媒-病毒-寄主植物三者之間的交互作用亦扮演不容忽視的角色。已 有研究報告指出,不管是昆蟲媒介動物病毒或植物病毒,這些病毒會改變其 病媒昆蟲適存值 (fitness) 的例子 (Sisterson, 2009)。以往的研究學者多認為 病毒對病媒昆蟲僅有負面影響,包含降低存活率、增長發育時間 (DeAngelis et al., 1993),但越來越多的研究指出除了負 面影響外尚有其他類 型的關係,
例如正面影響 (Bautista et al., 1995; Maris et al., 2004; Belliure et al., 2005;
St ump f a nd Ke nned y, 2007 ; Abe e t al. , 2012) 與無影響 (Wijkamp et al., 1996)。 據 Maris et al. (2004) 研 究 發 現 當 健 康 西 方 花 薊 馬 (Frankliniella occidentalis) 的 幼 蟲 飼 養 於 感 染 番 茄 斑 萎 病 毒 (Tomato spotted wilt virus, TSWV) 植 物 時 , 其 發 育 速 率 顯 著 高 於 飼 養 在 健 康 植 物 上 的 幼 蟲 。 至 於 TSWV 對 其 病 媒 薊 馬 存 活 率 之 影 響 方 面 , 將 西 方 花 薊 馬 成 蟲 飼 養 於 健 康 植 物 與 感 染 兩 種 不 同 分 離 株 TS WV 之 寄 主 植 物。結 果 發 現 飼 養 於 感 染 TSWV 之 CFL 分 離 株 植 物 上 的 薊 馬,其 幼 蟲 存 活 率 顯 著 高 於 飼 養 於 健 康 植 物 者;但 飼 養 於 感 染 TS WV 之 RG2 分 離 株 植 物 上 的 薊 馬 幼 蟲 , 其 存 活 率 與 飼 養 於 健 康 植 物 者 無 顯 著 差 異 (Stumpf and Kennedy, 2007)。 由 此 可 知 TS WV 分 離 株 的 不 同 , 對 病 媒 昆 蟲 造 成 的 效 應 亦 有 所 不 同。在 產 卵 量 方 面,據 Wijkamp et al. (1996) 以 矮 牽 牛 (Petunia x hybri da Ho rt ) 葉 片 供 健 康 以 及 感 染 TSWV 之 西 方 花 薊 馬 產 卵 , 結 果 發 現 其 產 卵 量 並 無 顯 著 差 異 。
有 關 薊 馬 對 不 同 感 染 狀 況 的 植 物 之 偏 好 性,Chaisuekul and Riley (2005) 指 出 煙 草 花 薊 馬 (F. fusca) 於 遭 TSWV 感 染 之 四 瓣 馬 齒 莧 (Portulaca oleraceae) 的產卵量顯著多於健康之四瓣馬齒莧,但在感染 TSWV 及健康番茄 植株的產卵量並沒有顯著差異。在 Maris et al. (2004) 則顯示 F. occidentalis 停留
3
於受 TSWV 感染之甜椒 (Capsicum annuum) 的蟲數顯著多於健康甜椒。這 顯 示 病 媒 薊 馬 對 寄 主 植 物 的 偏 好 性 會 隨 病 媒 薊 馬 的 種 類 、 寄 主 植 物 的 種 類 與 健 康 狀 況 而 有 所 不 同 。
雖 然 目 前 有 關 薊 馬 媒 介 植 物 病 毒 的 研 究 很 多 , 但研究最多的模式系 統為西方花薊馬與 TSWV,然 而 在 過 去 研 究 人 員 往 往 直 接 將 其 研 究 結 果 延 伸 至 其 他 種 類 的 薊 馬 及 其 他 薊 馬 媒 介 的 病 毒 。 可 是 以 薊 馬 幼 蟲 存 活 率 為 例 , 前 述 Stumpf and Kenned y (2007) 發 現 以 TSWV 對 西 方 花 薊 馬 幼 蟲 的 存 活 率 的 影 響 來 說 , 就 會 隨 不 同 病 毒 分 離 株 而 改 變 , 更 何 況 不 同 的 薊 馬 種 類 及 不 同 的 病 毒 種 類 。 此 外 在 臺 灣 西 方 花 薊 馬 及 TS WV 並 不 是 主 要 的 薊 馬 - 病 毒 系 統 , 故 本 研 究 採 用 發 生 於 臺 灣 地 區 瓜 類 作 物 上 重 要 的 病 媒 薊 馬 - 南 黃 薊 馬 及 其 傳 播 重 要 病 毒 - WS MoV 為 研 究 對 象 。 南 黃 薊 馬 廣 泛 分 布 於 世 界 各 地 , 包 括 歐 洲 、 亞 洲 、 非 洲 、 北 美 洲 、 南 美 洲 、 大 洋 洲 地 區 (Bhatti, 1998; Canno n et al., 2007), 其 寄 主 植 物 範 圍 相 當 廣 泛,可 為 害 多 種 蔬 菜、瓜 果 類 及 花 卉 (黃 及 陳,2004)。
南 黃 薊 馬 可 傳 播 超 過 4 種 以 上 的 Tospovirus 屬 的 病 毒 , 包 含 :彩色 海 芋 黃 化 斑 點 病 毒 (Calla lily chlorotic spot virus)、番椒黃化病毒 (Capsicum chlorosis virus)、洋香瓜黃斑病毒 (Melon yellow spot virus)、花生頂芽壞疽病 毒 (Groundnut bud necrosis virus)、西瓜頂芽壞疽病毒 (Watermelon bud necrosis virus), 重 要 的 是 此 薊 馬 乃 WS Mo V 唯 一 的 蟲 媒 薊 馬 種 類 (Ye h et al. , 1992 ; Pappu et al., 2009)。 病 毒 方 面 , W SMo V 為 臺 灣 地 區 第 一 種 被 發 現 的 Tospovirus 屬 的 病 毒 , 主 要 感 染 西 瓜 與 洋 香 瓜 等 多 種 葫 蘆 科 (Cucur bit a ce ae) 作 物,如 果 瓜 類 作 物 感 染 WS Mo V 將 對 產 量 造 成 重 大 損 失 (Yeh et al., 1992; 陳 等 2004)。
近年來有關南黃薊馬 及 WS MoV 的 研 究,大 多 著 重 南 黃 薊 馬 的 生 物 學 或 WS MoV 的 病 毒 學 研 究。媒 介 昆 蟲 方 面,多 針 對 南 黃 薊 馬 的 生 殖 與 發 育 進 行 探 討 (黃 及 蘇 , 1997; 黃 及 陳 , 2004); 病 毒 方 面 , 則 偏 重 於 WS MoV 本 身 性 質 研 究 , 如 對 其 L RNA、 M RNA、 S RNA 進 行 解 序 比 較 、 研 究 WSMoV 基 因 表 現 與 病 毒 血 清 群 (serogroup) 研 究
4
(Ye h et al., 1996 ; C hu a nd Ye h, 19 98 ; C hu et al., 2001 ; C he n et a l., 2010 ; Okuda et al., 2003)。 在 南 黃 薊 馬 傳 播 WSMo V 生 物 學 方 面 , 分 別 有 陳 等 (2004) 就 WSMoV 於 田 間 的 發 生 生 態 以 及 雜 草 寄 主 進 行 報 告 。 另 外 Chen et al. (2006) 亦 報 導 南 黃 薊 馬 可 能 可 以 非 持 續 性 或 持 續 性 兩 種 傳 播 模 式 傳 播 WS MoV。除 了 以 上 列 舉 的 報 告 外,尚 無 關 於 WSMoV 及 南 黃 薊 馬 之 間 的 交 互 作 用 報 導 。 因 此 , 確 定 南 黃 薊 馬 傳 播 WS MoV 傳 播 模 式 與 瞭 解 有 關 WS MoV 及 南 黃 薊 馬 之 間 交 互 作 用 有 其 必 要 性 。
本 研 究 目 的 有 二 : (1) 南 黃 薊 馬 傳 播 WSMoV 的 傳 播 生 態 學 , 包 含 南 黃 薊 馬 傳 播 病 毒 機 制 的 再 確 認 、 不 同 發 育 階 段 與 性 別 對 薊 馬 傳 播 病 毒 效 率 之 影 響;(2) 深 入 研 究 病 毒 與 病 媒 的 交 互 作 用,探 討 WS MoV 的 感 染 對 南 黃 薊 馬 幼 蟲 存 活 率 、 發 育 速 率 及 薊 馬 成 蟲 取 食 偏 好 性 的 影 響 。
5 材料與方法 1. 南黃薊馬、植物及病毒
南 黃 薊 馬 蟲 源 採 集 於 自 國 立 臺 灣 大 學 實 驗 農 場 的 茄 子 (Solanum melongena) 葉上,經形態及分子 檢測確認 (EPPO/CABI, 1997; 王,2002)。
南黃薊馬的 stock population 採用發芽 4 日的花豆苗 (Phaseolus coccineus) 飼養,將連同花盆的豆苗 置入直徑 7.5 公分,高 16 公分,底部鋪有珍珠 石的玻璃罐中 ,且罐口罩上每平方公分 250 網目的細紗網,於生長箱飼養 (25 ± 2℃,16L:8D),爾 後每 3 日更換一次新的花豆苗供薊馬取 食。為獲得 相 同 齡 期 之 供 試 南 黃 薊 馬 , 收 集 400-500 隻 南 黃 薊 馬 成 蟲 飼 養 於 小 黃 瓜 (Cucumis sativus) 上並提供花粉以促 進其產 卵。小黃瓜放置 於底部鋪有 擦手 紙的 2 公升燒杯中,且杯口罩上每平方公分 250 網目的細紗網,5 日後,
挑取孵化 4 小時內初生的一齡薊馬幼蟲供實驗用。
本實 驗 採用 的 WSMoV 台南 分 離株 (isolate) 係由台南 農業 改 良場 彭 瑞菊助理研究員提供; 採用 西瓜苗 (C. lanatus ‘F ine light ’) 維持實驗用 WSMoV,種植於生長箱 (25 ± 2℃,16L:8D)。實驗前將台南農業改 良場提 供 感染 WSMoV 的洋香瓜病葉以機械接種方式接種於圓葉菸草 (Nicotiana benthamiana),再將確認感 染 WSMoV 的菸草作為機械接種的病 源接種至僅 具一片真葉的西瓜苗 子葉上。機械接種方法概述如下:將欲接種的菸草葉面 灑布金剛砂 (carborudum powder, 500 mesh),取感染 WSMoV 之植物葉片 1g 加 於 裝 有 少 量 矽 藻 土 (celite) 及 10 ml 冰 冷 接 種 緩 衝 液 (0.01 M phosphate buffer, pH 7.0, 1% Na2SO3) 之冰冷研缽中研磨後,將 感染 WSMoV 的植物研磨液沾於衛生紙塗抹於接種葉面上,待 1 分鐘後以蒸餾水沖洗乾 淨。為確保 WSMoV 不因多次機械接種失去可被薊馬傳播的能力,WSMoV 平時以南黃薊馬接種 維持於西瓜苗上。待要實驗時,為獲得沒有遭到薊馬取食 損害的供試病源植物,先以上述的機械接種方法將 WSMoV 自西瓜接種至圓 葉菸草上,待圓葉菸草產生病徵時 (圖一B),再將 WSMoV 自圓葉菸草以 前述機械接種 方式接種至僅具一片真葉之西瓜苗 ,並給予 2 週時間發展病 勢 , 2 週 後 以 病 毒 專 一 性 引 子 對 進 行 反 轉 錄 聚 合 酶 連 鎖 反 應 (Reverse
6
transcription polymerase chain reaction, RT-PCR) 檢 測 西 瓜 苗 是 否 受 WSMoV 感染 (圖二B),以確定成功接種 病毒。
圖一、(A) 健康圓葉煙草 (Nicotiana benthamiana)。(B) 感染西瓜銀斑病毒 (Watermelon silver mottle virus, WSMoV) 之圓葉菸草。當圓葉煙 草感 染 WSMoV 時病徵為新葉捲曲皺縮。
Fig. 1. (A) Healthy tobacco (Nicotiana benthamiana). (B) The symptoms of Watermelon silver mottle virus-infected tobacco seedling are crinkling and curling of new foliors.
(A) (B)
7
圖 二 、 (A) 健 康 西 瓜 苗 (Citrullus lanatus) 。 (B) 感 染 西 瓜 銀 斑 病 毒 (Watermelon silver mottle virus, WSMoV) 之西瓜苗。當西 瓜苗感染 WSMoV 時病徵為新葉明顯黃化、皺縮叢 生、直立、呈壞疽 與焦枝 現 象。
Fig. 2. (A) Healthy watermelon (Citrullus lanatus). (B) The symptoms of Watermelon silver mottle virus-infected watermelon seedling are leaf mottling, crinkling, upright growth of younger branch, tip necrosis and dieback.
2. 西 瓜 銀 斑 病 毒 的 檢 測
所 有 的 病 源 植物 (source plant) 均 以 RT-PCR 檢 測 確 認 WSMoV 的 感染。RT-PCR 使用專一性引子對 WSMoV848 (5’-ATCACCATCATCCACA G-3’) 與 WSMoVR (5’-GAGGAGAGAGCAATC GAGG C-3'),乃修改自 Uga and Tsuda (2005),依據 WSMoV 之 S RNA 上的鞘蛋白基因 (Nucleocapsid protein gene) 及 3’ 非編碼區 (3’untranslated region) 設計。此引子對將增 幅出大小約為 800 bp 的核酸產物。以 TRIzol (Invitrogen, Carlsbad, CA) 抽 取供試植物的 total RNA,取 0.01 g 供試植物葉片,於 100 l TRIzol 與液 態氮一起研磨,其餘的相分離 (phase separation) 及 RNA 沉澱流程,均遵 照操作手冊操作。Total RNA 抽取完成後,遵照 One step RT-PCR kit (Qiagen,
(A) (B)
8
Valencia, CA) 操作流程,反應總 體積為 10 l。 將 1 l 植物 total RNA,
分 別 各 加 入 0.6 l 之 WSMoV848 與 WSMoVR 引 子 對 、 2 l 之 5X RT-PCR buffer、0.4 l 之 10 mM dNTP 與 0.4 l 之 Enzyme Mix, 其餘體 積以 DEPC 水補滿。並於 PCR 熱循環反應儀中 (PCR thermal cycler) 進 行反轉錄及 40 個 PCR 循環反應。RT-PCR 的反應條件如下:於 50℃ 30 分 鐘 的 條件 下 合 成 cDNA, 之 後 再 於 95℃ 進 行 變 性 (denature) 反 應 15 分鐘,再進行 40 個 PCR 循環反應,每一個循環反應之程序為 94℃ 變性 反應 30 秒,50℃ 黏合反應 (annealing) 30 秒,72℃ 聚合反應 (elongation) 1 分鐘,最後再於 72℃ 進行延長 (extension) 聚合反應 10 分鐘反應結果 以 1% 瓊脂膠體 (agarose) 電泳進行分析,預期增幅出核酸產物的大小 約 為 800 bp。
南 黃薊 馬 傳播 WSMoV 傳播模式實驗中,供試植物 WSMoV 感染檢 測 部 分 , 採用 total RNA 粗萃取進行 RT-PCR。在實驗前進行前測試確認 toatal RNA 粗萃取進行 RT-PCR 與 TRIzol 抽取 total RNA 進行 RT-PCR 檢 測 效 率 完全 相同 。 total RNA 粗萃取方 法敘 述如下:將待測葉片以 1:20 (w/v) 於 ELISA grinding buffer (1.59 g/l Na2CO3, 2.93 g/l NaHCO3, pH 9.6, 2% PVP-40, 0.2% bovine serum albumin, 0.05% Tween 20 及 1% Na2S2O5) 研磨。取 2 l 研磨液加入 25 l GES denaturing buffer 中 (0.1 M glycine, 0.05 M NaCl, 1 mM EDTA 及 0.5% Triton X-100)。於 95℃ 加熱 10 分鐘 後 , 迅 速 降 溫 至 4℃ 維 持 10 分 鐘 。 將 1 l 前 述 GES 均 質 液 (homogenate) 作為 RT-PCR 的 RNA 模板,進行 RT-PCR。其餘 RT-PCR 操作流程皆同前述。
3. 南黃薊馬傳播西瓜銀斑病毒的傳播模 式
為 確認 南黃 薊馬 以何 種 傳播模 式 傳 播 WSMoV,以及 南黃薊馬獲得病 毒齡期 對病 毒 傳 播 效率 的 影 響。 將指 定齡 期的薊 馬幼 蟲 (孵化時 間 小 於 4 小 時 之 一 齡幼蟲 、 蛻皮小 於 4 小時之二齡幼蟲 ) 或成蟲 (48 小時內羽化) 以沾水的水彩筆挑取,置於經 RT-PCR 檢測確認感染 WSMoV 的西瓜葉片
9
上, 分別 給予 指定 的 AAP,待指定 AAP 到達時將 薊馬轉移至僅具一片真 葉之西瓜植株上進行 接種停留時間 (inoculation access period, IAP)。在持續 性傳播模式測試中,指定 AAP 結束後,則會先將薊馬轉移至花豆葉上給予 指定的潛伏期,待潛伏期過後將薊馬轉移至供試西瓜植株進行 IAP。傳播模 式確認實驗中,每株 實驗植株上進行 IAP 的薊馬蟲數,除了持續性傳播模 式一齡幼蟲獲 毒成蟲傳毒的部分 ,為了區分不同性別薊馬傳播 WSMoV 之 傳播效率,每株供試 植物各接種 1 隻薊馬成蟲外;其餘傳播模式測試為了 提高接種病毒感染的成功率,每株供試植 物各接種 5 隻薊馬。在指定的 IAP 結束後,噴灑稀釋 1000 倍之 20% 亞滅培 (Acetamiprid) 以除去供試植物 上的薊馬,並將供試植株移入生長箱待其病勢發展。供試植株移入生長箱 2 週後,以前述粗萃取方法抽取供試植物葉片 的 total RNA,以 RT-PCR 檢測 是否有 WSMoV 感染,並 計算南黃薊馬傳播 WSMoV 之傳播效率。此外,
在傳播模式測試中作為獲毒來源的染病西瓜葉片在薊馬 AAP 結束後,為確 認 薊 馬 獲 毒 來 源 確 實 有 WSMoV 的 感 染 , 所 有 葉 片 均 以 TRIzol 抽 取 其 total RNA, 再以 RT-PCR 進行病毒感染檢 測。每種傳播模式測試,各有 3 組實驗組,每組均有 10 株供試植物。所有傳播模式確認實驗都有進行生長 箱負對照組試驗。生長箱負對照組 試驗內容如下:隨機選取與傳播實驗相同 批次且未被薊馬取食 過僅具一片真葉的 健 康西瓜苗 5 株,與薊馬接種後之 供試植物一同 放置於生長箱內 2 週。在生長箱負對照組試驗中並沒有西瓜 植株被檢測出有 WSMoV 感染。詳細的各傳播模式的試驗條件,敘述如下:
3.1 非持續性傳播模式 3.1.1 一齡幼蟲獲毒及傳毒
將 薊 馬 一 齡 幼 蟲 置 於 確 認 感 染 WSMoV 的 西 瓜 葉 片 上 15 分 鐘 (AAP),然後 將 可能 帶 毒 的 薊 馬 每 5 隻 轉 移 至 僅 具 一 片真葉 的 健康西瓜苗 上進行接種 2 小時 (IAP)。
10 3.1.2 二齡幼蟲獲毒及傳毒
將薊馬二齡幼蟲經 1 小時飢餓期後,置於確認感染 WSMoV 的西瓜葉 片上 15 分鐘 (AAP),然後將可能獲毒的薊馬每 5 隻轉移至僅具一片真葉 之健康西瓜苗上,進行接種 2 小時 (IAP)。
3.1.3 成蟲獲毒及傳毒
將薊馬成蟲經 1 小時飢餓期後,置於確認感染 WSMoV 的西瓜葉片上 15 分鐘 (AAP), 然後 將 可 能 獲 毒 的 薊 馬每 5 隻 轉移 至 僅具 一片真葉之 健 康西瓜苗上進行接種 2 小時 (IAP)。
3.2 半持續性傳播模式 3.2.1 一齡幼蟲獲毒及傳毒
將 薊 馬 一 齡 幼 蟲 , 放 置 於 確 認 感 染 WSMoV 的 西 瓜 葉 片 上 2 小 時 (AAP),其後將 可能 獲 毒 的 薊 馬 每 5 隻 轉 移 至 僅 具一片真葉 之 健康 西瓜 苗 上進行接種 24 小時 (IAP)。
3.2.2 二齡幼蟲獲毒及傳毒
將 薊 馬 二 齡 幼 蟲 , 放 置 於 確 認 感 染 WSMoV 的 西 瓜 葉 片 上 2 小 時 (AAP),其後將 可能 獲 毒 的 薊 馬 每 5 隻 轉 移 至 僅 具一片真葉 之 健康 西瓜苗 上,進行接種 24 小時 (IAP)。
3.2.3 成蟲獲毒及傳毒
將薊馬成蟲,放置於 確認感染 WSMoV 的西瓜葉片上 2 小時 (AAP),
其後將 可能獲 毒的薊 馬每 5 隻轉移至僅具一片真葉之健康西瓜苗上,進行 接種 48 小時 (IAP)。
11 3.3 持續性傳播模式
持續性增殖型傳播模式測試,依照薊馬獲毒齡期不同,分為一齡幼蟲獲 毒、二 齡幼蟲獲毒與成蟲 獲毒。當薊馬於確認感染 WSMoV 的西瓜葉片結 束 AAP 後,將薊馬轉移至花豆葉上給予適當潛伏期,待潛伏期後將薊馬幼 蟲再轉移至僅具一片真葉之西瓜苗 進行 IAP。其中,薊馬一齡幼蟲獲毒部分 又可依傳毒齡期不同分為二齡幼蟲傳毒及 成蟲期傳毒。以下為詳細流程:
3.3.1 一齡幼蟲獲毒、成蟲傳毒
將 薊 馬 一 齡 幼 蟲 置 於 確 認 感 染 WSMoV 的 西 瓜 葉 片 上 取 食 48 小 時 (AAP) , 然 後 將 可 能 獲 得 病 毒 的 薊 馬 幼 蟲 轉 移 至 置 有 (1.5 X 1.5 cm2) 非 WSMoV 寄主植物花豆葉 (leaf disk) 之玻 璃瓶直徑 1.5 公分 高 4.5 公分 中飼養,並以二層石蠟膜 (parafilm) 封住瓶口。飼養至成蟲期後,將可能帶 毒 的 薊 馬 成 蟲 單 隻 轉 移 至 僅 具 有 一 片 真 葉 之 健 康 西 瓜 苗 接 種 48 小 時 (IAP)。為比較不同性別薊馬傳 播 WSMoV 效率,每組 10 株植株,每株植 株各放置 1 隻薊馬成蟲進行 IAP。
3.3.2 一齡幼蟲獲毒、二齡幼蟲傳毒
將 薊 馬 一 齡 幼 蟲 置 於 確 認 感 染 WSMoV 的 西 瓜 葉 上 給 予 48 小 時 (AAP) 後,將其 以花豆葉飼養 至二齡幼蟲。將 可 能帶毒二齡幼蟲,每 5 隻 轉移置於僅具一片真葉之健康西瓜苗上給 予 24 小時的取食時間 (IAP)。
3.3.3 二齡幼蟲獲毒、成蟲傳毒
將 薊 馬 二 齡 幼 蟲 置 於 確 認 感 染 WSMoV 的 西 瓜 葉 上 取 食 48 小 時 (AAP) 後。以前述的方法飼養至成蟲,當成 蟲羽化後,再將可能帶毒成蟲 每 5 隻 轉 移 至 僅 具 一 片 真 葉 之 健 康 西 瓜 苗 上 給 予 48 小 時 的 取 食 時 間 (IAP)。
12 3.3.4 成蟲獲毒、成蟲傳毒
將 薊 馬 成 蟲 置 於 確 認 感 染 WSMoV 的 西 瓜 葉 片 上 取 食 48 小 時 (AAP),然後轉 移可能 獲毒 薊 馬 ,每 5 隻 轉移至 僅具 一片真 葉之 健 康 西瓜 苗上給予 48 小時的取食時間 (IAP)。
4. 西瓜銀斑病毒於南黃薊馬體內的複製
將經前述 RT-PCR 檢測確認感染 WSMoV 的薊馬之 total RNA,以針 對 WSMoV M RNA 之 viral complementary RNA (vc RNA) 設計之引子 NSmR (5’-GCTGTGTAGCTGAGCGCAA-3’) 進行反轉錄產生 cDNA 後,再 以 引 子 對 NSmF (5’-AATTTTAGTGATCATGGCTGTG-3’) 及 NSmR 對 反 轉錄產生之 cDNA 進行 PCR 增幅,以確認 WSMoV 是否於病媒薊馬體內 進 行 複 製 。薊 馬 的 total RNA 以 TRIzol 抽取,取 1 隻薊馬,於 100 l TRIzol 與液態氮研磨,其餘操作程序同 上 述。所 抽取出的薊馬 total RNA 以 ReverTra Ace (TOYOBO, Osaka, Japan) 進行反轉錄。反轉錄反 應總體積 為 20 l,加入 1 l 之薊馬 total RNA、4 l 之 5X Reaction buffer 及 1 l 之引子 NSmR、0.25 l 之 RNase inhibitor、2 l 之 10 mM dNTP、1 l 之 ReverTra Ace,
其餘體積以 DEPC 水補滿,於 PCR 熱循環反應儀進行反轉錄, 反轉錄條 件如下,42℃ 20 分鐘,99℃ 5 分鐘,4℃ 5 分鐘。將前述的 cDNA 以 2X PCR Mastermix RED (Bioman, Taipei, Taiwan) 進行 PCR 增幅,PCR 反應總 體積為 20 l,加入 2 l 之 cDNA,10 l 之 2X Mastermix Red,各 1.2 l 引 子 對 NSmF 及 NSmR, 其餘體積以無菌 雙蒸餾水補滿, 反應條件如下,
於 95℃ 變性反應 2 分鐘,再進行 35 個 PCR 循環反應,每一個循環反 應 之 程 序 為 95℃ 變性反應 30 秒,55℃ 黏合反應 30 秒,72℃ 聚合反 應 1 分鐘,最後再於 72℃ 進行延長聚合反應 10 分鐘,反應結果以 1%
瓊 脂 膠 體電 泳 進 行分 析 ,預 期 增幅 出 的 WSMoV M RNA 之 vc RNA 的 RT-PCR 產物大小約為 275 bp。
13
5. 西瓜銀斑病毒對南黃薊馬幼蟲發育速 率 及存活率影響
為了檢驗 WSMoV 對南黃薊馬幼蟲的發育速率及存活率影響,分別使 用健康薊馬幼蟲及感染 WSMoV 薊馬幼蟲進行試驗。感染 WSMoV 薊馬幼 蟲 部 分 , 將 孵 化 4 小 時 內 的 薊 馬 幼 蟲 , 放 置 於 RT-PCR 檢 測 確 認 感 染 WSMoV 西瓜葉片上給予 24 小時的 AAP。AAP 結束後將前述可能帶 毒的 薊馬幼蟲 (帶毒率 90%) 單隻飼養於內置有 1 X 1 cm2 之花豆葉片或西瓜 葉片培養皿 (直徑 3.5 cm),以石蠟膜密封 。每 12 小時記錄薊馬幼蟲發育 階段,直到前蛹期 (prepupa stage)。最後統計幼蟲發育速率及存活率。花豆 或西瓜葉片視葉片情 況,每 24~48 小時更換一次。每次實驗蟲數介於 30~40 隻不等。健康薊馬部分,除了前 24 小時 AAP 取食葉片為健康西瓜葉片外,
其餘處理均與感染 WSMoV 的薊馬相同。
5.1 直接效應
為 調 查 WSMoV 的 感 染 對 南 黃 薊 馬 幼 蟲 發 育 速 率 及 幼 蟲 存 活 率 影 響 的直接 影響,採用不會被 WSMoV 感染之非寄主植物花豆,做為薊馬一齡 幼蟲到前蛹期食物。分別比較感染 WSMoV 的薊馬幼蟲與未受 WSMoV 感 染的薊馬幼蟲於花豆葉上之發育速率及存活率。
5.2 間接效應
為研 究薊馬取食感染 WSMoV 的植物對薊馬生存及發育 所受的影響,
分別讓健康薊馬取食 健康西瓜葉片、經薊馬 取食的西瓜葉片與經薊馬接種感 染 WSMoV 的西瓜葉片;感染 WSMoV 的薊馬幼蟲取食健康 西瓜葉片、經 薊馬取 食的西瓜葉片與經 薊馬接種感 染 WSMoV 的西瓜葉片。 觀察記錄健 康薊馬及感染 WSMoV 的薊馬自一齡幼蟲至前蛹期之發育時間 及存活率。
14 6. 南黃薊馬成蟲之取食偏好性
設置 1 個 30 x 30 x 30 cm3 壓克力箱,內 置入兩株不同實驗組合條件 植物 (健康植株與薊馬接種 感染 WSMoV 之植株、健康植株與經薊馬取食 植株、經薊馬取食植株與薊馬接種感染 WSMoV 之植株),並將內有同樣性 別 10 隻薊馬成蟲的離心管 (eppendorf) 放置於 2 植株中央;植株與植株、
各植株與 離心 管之 間隔均為 30 公分。待放置 2 小時、4 小時、8 小時、
12 小時及 24 小時,後分別計 算於各 植株 上之薊馬 數量。 本實驗 每個實驗 處理均進行 3 次重覆,另外雌薊馬與雄薊馬都分別進行測試,以檢測薊馬 成蟲取食偏好性之性別差異。
7. 統計分析
南黃薊馬 傳播 WSMoV 之傳播模式檢測部分,單隻薊馬傳播 WSMoV 之傳播效率計算乃根據以下 Swallow (1985) 的群組傳播效率計算公式計算,
其中 N 為供試植物總數,R 為沒有感染成功的植物數,k 為每株供試植物 接種的薊馬數:PS = 1-﹛ R/N﹜1/k。在持續性傳播測試一齡幼蟲獲毒、成蟲傳 毒 的 實 驗 中, 其單 隻 薊馬 成蟲 傳 播 病毒的效率為 感染植 物總數/供試總植物 數 。 不同性別成蟲傳播效率的比較以邏輯迴歸 (logistic regression) 進行分 析 。 在 WSMoV 對 薊 馬 幼 蟲 的 存 活 率 直 接 、 間 接 的 影 響 , 本 研 究 採 用 Kaplan-Meier 法 及 存 活 分 析 (survival analysis) 檢 驗 薊 馬 幼 蟲 存 活 率 (cumulative survival rate) 或 存 活 至 前 蛹 期 的 總 蟲 數 。 利 用 對 數 等 級 檢 定 (log rank test) 檢視兩處理之間是否具有顯 著差異。 WSMoV 對薊馬幼蟲發 育時間的直接及間接效應 以負二項分布 (negative binomial distribution) 進 行比較。 薊馬成蟲對 薊馬接種感染 WSMoV 植物、經薊馬取食 植物與健康 植物取食偏好性測試,每個時間點兩植株上薊馬蟲數比較利用,以二項分佈 (binomial distribution) 為基礎之符號檢定 (sign test) 進行檢定。以上所有的 統計運算均由軟體 R (ver. 2.12.0) 執行。
15 結果 1. 南黃薊馬傳播西瓜銀斑病毒的傳播模 式 1.1 非持續性傳播模式
南 黃 薊 馬 無 法 在 15 分 鐘 AAP 與 2 小 時 IAP 的 情 況 下 傳 播 WSMoV。在 本實 驗中 一齡 幼蟲 獲毒 及傳 毒 、二齡 幼蟲 獲毒 及傳 毒、 成蟲 獲 毒及傳毒,單隻薊馬傳播 WSMoV 的效率皆為 0% (表一)。
1.2 半持續性傳播模式
南 黃 薊 馬 雖 然 可 以 在 2 小 時 AAP 與 24 小 時 IAP 的 情 況 下 傳 播 WSMoV,但 是傳 播機 率非 常低 。 在 本實 驗 中一齡 幼蟲 獲毒 及傳 毒、 二齡 幼 蟲獲毒及傳毒、成蟲獲毒及傳 毒,5 隻薊馬傳播 WSMoV 的效率分別為 3%、
6%、0% (表一),一齡 與二齡幼蟲的傳 播效 率並沒有顯著差異 (z = -0.55, d. f.
= 1, P = 0.57)。
1.3 持續性傳播模式
南 黃薊 馬 主要 以 持續 性 傳播模 式 傳播 WSMoV,其中以一齡幼蟲獲毒 成蟲傳毒的傳播效率最高。本實驗中一齡幼蟲及二齡幼蟲獲毒,且於羽化為 成蟲後傳播病毒,單隻薊馬的 傳播效率分別為 76% 及 0.6% (表一)。一齡 幼蟲獲毒二齡幼蟲傳毒部分,單隻薊馬的傳播效率為 0.6% (表一)。成蟲獲 毒及傳毒部分,單隻薊馬的傳播效率為 1.2% (表一)。雌成蟲與雄成蟲傳播 WSMoV 的傳播效率,分別為 71% 及 81% ,兩者間無顯著差異 (z = -0.23, d. f. = 1, P = 0.81)。
16
表一、南黃薊馬 (Thrips palmi) 傳播西瓜銀斑病毒 (Watermelon silver mottle virus, WSMoV) 之傳播效率
Table 1. Transmission efficiency of Watermelon silver mottle virus (WSMoV) by Thrips palmi
Transmission mode
Acquired instar Inoculated instar Transmission efficiencya
Psb
Nonpersistent
transmission First-instar larvae First-instar larvae 0% 0%
Second-instar larvae Second-instar larvae 0% 0%
Adults Adults 0% 0%
Semipersistent
transmission First-instar larvae First-instar larvae 3% 0.6%
Second-instar larvae Second-instar larvae 6% 1.2%
Adults Adults 0% 0%
Persistent
transmission First-instar larvae Second-instar larvae 3% 0.6%
Second-instar larvae Adults 76% 76%
Second-instar larvae Adults 3% 0.6%
Adults Adults 6% 1.2%
a Transmission efficiency cauculated as the number of WSMoV-infected plants/
the number of test plants.
b The maximum likelihood estimator of transm ission probability for replicates with five thrips per plant was counted according to Swallow (1985).
17 2. 西瓜銀斑病毒於南黃薊馬體內的複製
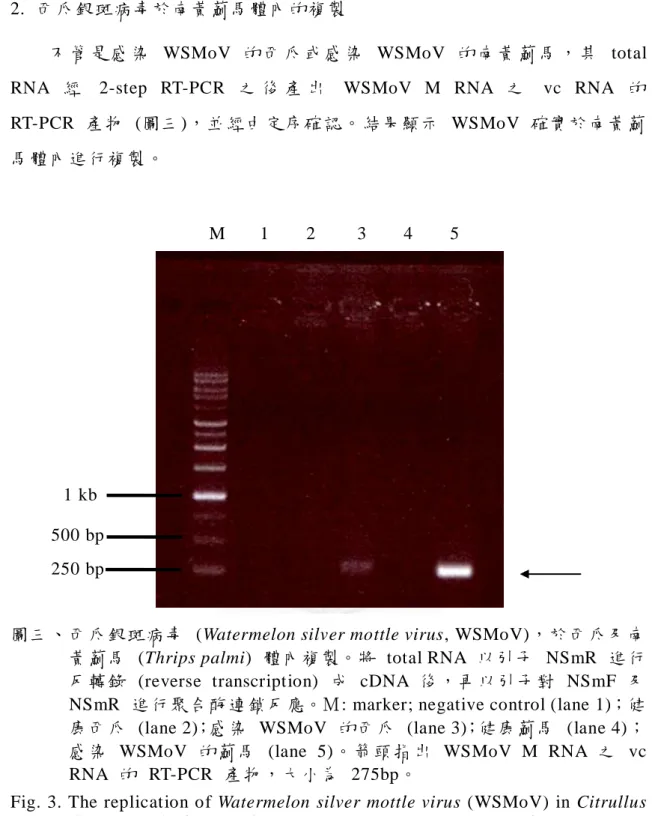
不 管 是 感 染 WSMoV 的 西 瓜 或 感 染 WSMoV 的 南 黃 薊 馬 , 其 total RNA 經 2-step RT-PCR 之 後 產 出 WSMoV M RNA 之 vc RNA 的 RT-PCR 產物 (圖三),並經由定序確 認。 結果顯示 WSMoV 確實於南黃薊 馬體內進行複製。
圖三、西瓜銀斑病毒 (Watermelon silver mottle virus, WSMoV),於西瓜及南 黃薊馬 (Thrips palmi) 體內複製。將 total RNA 以引子 NSmR 進行 反轉錄 (reverse transcription) 成 cDNA 後,再以引子對 NSmF 及 NSmR 進行聚合酶 連鎖反應。M: marker; negative control (lane 1);健 康西瓜 (lane 2);感染 WSMoV 的西瓜 (lane 3);健康薊馬 (lane 4);
感染 WSMoV 的 薊馬 (lane 5)。箭頭指出 WSMoV M RNA 之 vc RNA 的 RT-PCR 產物,大小為 275bp。
Fig. 3. The replication of Watermelon silver mottle virus (WSMoV) in Citrullus lanatus and Thrips palmi. Total RNA was reverse transcripted to cDNA by using the primer NSmR, and then the polymerase chain reaction was performed by using the cDNA and the primers NSmF and NSmR. M:
marker; negative control (lane 1); hea lthy watermelon (lane 2);
WSMoV-infected watermelon (lane 3); healthy thrips (lane 4);
WSMoV-infected thrips (lane 5). The arrow indicated the RT-PCR product (275bp) of WSMoV M RNA vc RNA.
M 1 2 3 4 5
1 kb 500 bp 250 bp
18
3. 西瓜銀斑病毒對南黃薊馬幼蟲發育速 率 及存活率之影響 3.1 直接效應
比較健康及感染 WSMoV 的南黃薊馬飼養於花豆上的存活率 (圖四),
兩者間並無顯著差異 (2 = 2.84, d. f. = 1, P = 0.09)。健康薊馬與感染 WSMoV 的薊馬於花豆上發育至前蛹期的時間分別為 119.4 ± 31.6 小時及 122.9 ± 18.4 小時,兩者間無顯著差異 (表二,Log-likelihood: -339.09, P = 0.73)。以 上結果顯示 WSMoV 的感染對薊馬幼蟲的存活率及發育至前蛹期的 時間無 直接影響。
表 二 、 西 瓜 銀 斑 病 毒 (Watermelon silver mottle virus) 的 感 染 對 南 黃 薊 馬 (Thrips palmi) 於花豆苗上的幼蟲 存活率與 幼蟲發育至前 蛹期時間 的 直接效應
Table 2. Direct effects of Watermelon silver mottle virus infection on larval survival rate and developmental time from newly larvae hatched to prepupae of Thrips palmi on kidney bean seedlings
Test statistic P-value
Survival rate 2.81a 0.09
Developmental time -339.09b 0.73
a The log-rank test was used for planned comparison. d. f. = 1.
b The negative binomial distribution was used for planned co mparision.
19
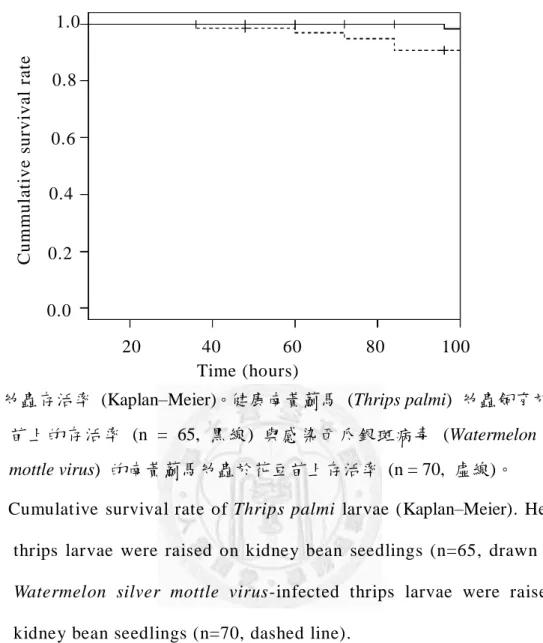
圖四、幼蟲存活率 (Kaplan–Meier)。健康南黃薊馬 (Thrips palmi) 幼蟲飼育於花豆 苗上的存活率 (n = 65, 黑線) 與感染西瓜銀斑病毒 (Watermelon silver mottle virus) 的南黃薊馬幼蟲於花豆苗上存活率 (n = 70, 虛線)。
Fig. 4. Cumulative survival rate of Thrips palmi larvae (Kaplan–Meier). Healthy thrips larvae were raised on kidney bean seedlings (n=65, drawn line).
Watermelon silver mottle virus-infected thrips larvae were raised on
kidney bean seedlings (n=70, dashed line).
3.2 間接效應
健 康南 黃薊 馬 幼蟲 飼 育 於 健康植 株 、 經 薊 馬 取 食植株及薊馬接種 感染 WSMoV 的植株之幼蟲存活率,三者 之間 沒有顯著差異 (表二、圖五 A,2= 4.6, d. f. = 2, P = 0.31)。結果顯示薊馬取食與 WSMoV 並不會透過薊馬的寄主 植物間接影響薊馬的存活率。發育至前蛹期時間方面,健康南黃薊馬幼蟲於 健康植株及薊馬取食 植株發育時間有顯著差異 (表三、圖六,Log-likelihood:
-235.10, P < 0.05);但健康南黃薊馬幼蟲於薊馬接種感染 WSMoV 之植株與 經薊馬取食之植株的發育時間,沒有顯著差異 (Log-likelihood: -163.44, P = 0.52);健康南 黃薊 馬幼蟲於 健康 植株 與薊 馬接種 感染 WSMoV 的植株發育
Time (hours)
20 40 60 80 100
0.0 0.2 0.4 0.6 0.8 1.0
Cummulative survival rate
20
時間亦無顯著差異 (Log-likelihood: -220.80, P = 0.53)。結果顯示 WSMoV 並 不會透過薊馬的寄主植物間接影響,薊馬的存活率與發育時間。
感染 WSMoV 之南黃薊馬幼蟲於健康植株、經薊馬取食植株及薊馬接 種感染 WSMoV 的植株之存活率,三者之間沒有顯著差異 (圖五B,2= 3.5, d. f. = 2, P = 0.17)。經薊馬取食與 WSMoV 對感染 WSMoV 之薊馬的存活率 沒有間接效應,此結果與健康薊馬的結果相同。
21
圖五、幼蟲存活率 (Kaplan–Meier)。(A) 健康南黃薊馬 (Thrips palmi) 幼蟲飼育於 健康西瓜苗上的存活率 (n = 47, 黑線),於薊馬取食西瓜苗上的存活率 (n = 32, 虛線),於薊馬接種感染西瓜銀斑病毒 (Watermelon silver mottle virus, WSMoV) 西瓜苗上的存活率 (n = 32, 灰線)。(B) 感染 WSMoV 之南黃薊 馬幼蟲飼育於健康西瓜苗上的存活率 (n = 46, 黑線),於薊馬取食西瓜苗上 的存活率 (n = 26, 虛線) ,於薊馬接種感染 WSMoV 西瓜苗上的存活率 (n = 40, 灰線)。
Fig. 5. Cumulative survival rate of Thrips palmi larvae (Kaplan–Meier). (A) Healthy thrips larvae were raised on healthy watermelon seedlings (n=47, drawn line), on thrips-damaged watermelon seedlings (n=32, dashed line) and on thrips-inoculated watermelon seedlings (n=32, grey line). (B) Watermelon silver mottle virus-infected thrips larvae were raised on healthy watermelon seedlings (n=46, drawn line), on thrips-damaged watermelon seedlings (n=26, dashed line) and on thrips-inoculated watermelon seedlings (n=40, grey line).
Time (hours)
(A) (B)
0.0 0.2 0.4 0.6 0.8 1.0
20 40 60 80 100 20 40 60 80 100
0 0
Cummulative survival rate
Time (hours)
22
發育至前蛹期時間方面,感染 WSMoV 的薊馬幼蟲於健康植株與薊馬 取食植 株之發育速 率有顯著差異 (表三、圖六,log-likelihood: 193.58, P <
0.01); 感 染 WSMoV 的 薊 馬幼 蟲 於 薊 馬 接 種 感 染 WSMoV 的 植 株及健康 植 株 之 發 育 速 率 有 顯 著 差 異 (log-likelihood: -266.85, P < 0.01) ; 感 染 WSMoV 的薊馬幼蟲於 薊馬接種 感染 WSMoV 的植株及經薊馬 取食植株之 發育速率有顯著差異 (log-likelihood: -193.58, P < 0.01)。感染 WSMoV 的 薊 馬 幼 蟲 於健 康植 株上發 育至 前蛹期 的 時 間最長 ,其次 為 於 薊馬接 種感 染 WSMoV 者,最短者為於經薊 馬取食植株發 育者。結 果顯示不論薊 馬有無感 染 WSMoV,當薊馬幼蟲飼育於 經薊馬取食的植株上會縮短薊馬幼蟲發育至 前蛹的發育時間,但 WSMoV 的感染會減弱薊馬取食植株造成的影響。
表三、西瓜銀斑病毒 (Watermelon silver mottle virus, WSMoV) 對南黃薊馬 (Thrips palmi) 於西瓜苗上的幼蟲發育至前 蛹期時間的 間接效應 Table 3. Indirect effects of Watermelon silver mottle virus (WSMoV) on larva
developmental time from newly hatched to prepupa of Thrips palmi on watermelon seedlings
Comparision
Healthy larvae WSMoV-infected larvae Test statistica P Test statistica P
Healthy vs. thrips-damaged -235.10 < 0.05 -203.92 < 0.05
Thrips-inoculated vs.
thrips-damaged
-163.44 0.52 -193.58 < 0.01
Healthy vs. thrips-inoculated -220.80 0.53 -266.85 < 0.01
a. The negative binomial distribution was used for planned comparision.
23
圖 六 、南 黃薊馬 (Thrips palmi) 幼蟲飼育 於西瓜葉 至前蛹期 的 平均 發育時 間 (平 均 ± 標 準 差 ) 。 薊馬幼蟲於薊馬接種感染西瓜銀斑病毒 (Watermelon silver mottle virus, WSMoV) 之西瓜葉發育時間 (黑色條帶,健康薊馬 n = 16;感染 WSMoV 之薊馬 n = 28);薊馬幼蟲於薊馬取食之西瓜葉發育時間 (灰色條帶,健康薊馬 n = 20;感染 WSMoV 之薊馬 n = 15);薊馬幼蟲於健 康之西瓜葉發育時間 (白色條帶,健康薊馬 n = 32;感染 WSMoV 之薊馬 n
= 28)。誤差線 (error bars) 表示標準差。圖中不同的字母表示數值間有顯著差 異 (經負二項分布比較之 P < 0.05)。
Fig. 6. Developmental time from newly hatcheded larva to prepupa of Thrips palmi on watermelon leaf. Black columns: healthy thrips on thrips-inoculated plants (number of prepupa = 16); Watermelon silver mottle virus (WSMoV)-infected thrips on thrips-inoculated plants (number of prepupa = 28); Grey columns: healthy thrips on thrips-damaged plants (number of prepupa = 20); WSMoV-infected thrips on thrips-damaged plants (number of prepupa = 15); White columns: healthy thrips on healthy plants (number of prepupa = 32); WSMoV-infected thrips on healthy plants (number of prepupa = 28). Different letters indicate statistical differences between means at P < 0.05 analyzed by negative binomial distribution.
0 50 100 150
Developmental time (hours)
Healthy thrips WSMoV-infected thrips ab
b
a a
c b
24 4. 南黃薊馬成蟲之取食偏好性
將 南黃 薊馬 成 蟲釋放於 健康植株 及 經 薊 馬 取食 的植 株 之間, 雌薊 馬在 2、8、12、24 或雄薊馬在釋放 2、12、24 小時均對薊馬取食植 物有較高的 偏好性 (圖七)。當南黃薊馬釋放於經薊馬取食及薊馬接種感染 WSMoV 的 植株間,雌薊馬在任何時間點對兩種植物的偏好性沒有差異,雄薊馬於釋放 2、 4 小 時 對薊 馬 接 種 感染 WSMoV 的 植株 有 較 高 的 偏好 性 (圖 八)。 這指 出 WSMoV 的感染可增加雄薊馬對植株的偏好度。當 南黃薊馬 釋放於健康 植株及薊馬接種感染 WSMoV 的植株間,雄薊馬在釋放後 4、8、12、24 小 時或雌薊馬在任何時間點對薊馬接種感染 WSMoV 的植株有較高的偏好性 (圖九)。結果顯示南黃薊馬偏 好取食感染 WSMoV 的植株。
25
圖七、南黃薊馬 (Thrips palmi) 成蟲對薊馬取食西瓜及健康西瓜植株的取食 偏好性。(A) 薊馬雄成蟲在三次偏好性選擇實驗中,停留於薊馬取食 西瓜 (灰色長條) 及健康西瓜 (白色長條) 植株上薊馬數量。(B) 薊馬 雌成蟲在 三 次偏好 性 選擇實 驗 中, 停 留 於薊馬取食 西 瓜 (灰色長條) 及健康西瓜 (白色長條) 植株上薊馬數量。每次實驗重覆在兩植物間釋放 10 隻薊馬成蟲。在偏好性實驗中 * 表示經符號檢定之 P < 0.05 以及 **
表示 P < 0.01。誤差線 (error bars) 表示標準差 (standard error)。
Fig. 7. Feeding preference of Thrips palmi adults to thrips-damaged versus healthy watermelon seedlings. (A) Number of male thrips adults on thrips-damaged (grey columns) and healthy seedlings (white columns) in three trials. (B) Number of female thrips adults on thrips-damaged (grey columns) and healthy seedlings (white columns) in three trials.
Ten adults were released between the two seedlings at the beginning of the experiment. * and ** indicate significant differences (P < 0.05 and P < 0.01) between numbers of thrips adults on thrips-damaged and healthy seedlings by sign test. Error bars indicate the standard error of the mean.
0 2 4 6 8
2 4 8 12 24
0 2 4 6 8
2 4 8 12 24
Number of thrips
Time after thrips release (hours)
(A) (B)
*
**
** **
**
**
**
Time after thrips release (hours)
26
圖八、南黃薊馬 (Thrips palmi) 成蟲對薊馬取食西瓜及薊馬接種感染西瓜銀 斑病毒 (Watermelon silver mottle virus, WSMoV) 之西瓜植株的取食 偏好性。(A) 薊馬雄成蟲在三次偏好性選擇實驗中,停留於薊馬取食 西瓜 (灰色 長條 ) 及 薊馬 接 種感 染 WSMoV 之西 瓜 (黑 色長 條 ) 植 株上薊馬數量。(B) 薊馬雌成蟲在三次偏好性選擇實驗中,停留於薊馬 取食西瓜 (灰色長條) 及薊馬接種感染 WSMoV 之西瓜 (黑色長條) 植株上薊馬數量。每次實驗重覆在兩植物間釋放 10 隻薊馬成蟲。在偏好 性實驗中 * 表示經符號檢定之 P < 0.05 以及 ** 表示 P < 0.01。誤差線 (error bars) 表示標準差 (standard error)。
Fig. 8. Feeding preference of Thrips palmi adults to thrips-damaged versus Watermelon silver mottle virus (WSMoV)-infected watermelon seedlings. (A) Number of male thrips adults on thrips-damaged (grey columns) and WSMoV-infected seedlings (black columns) in three trails. (B) Number of female thrips adults on thrips-damaged (grey columns) and WSMoV-infected seedlings (black columns) in three trials. Ten adults were released between the two plants at the beginning of the experiment. * and
** indicate significant differences (P < 0.05 and P < 0.01) between numbers of thrips adults on thrips-damaged and WSMoV-infected seedlings by sign test.
Error bars indicate the standard error of the mean.
0 2 4 6
2 4 8 12 24
0 2 4 6
2 4 8 12 24
(A) (B)
**
*
Number of thrips
Time after thrips release (hours) Time after thrips release (hours)
27
圖 九 、南黃 薊馬 (Thrips palmi) 成蟲對薊 馬接種感染 西瓜銀斑 病毒 (Watermelon silver mottle virus, WSMoV) 之西瓜及健康西瓜植株的取食偏好 性。(A) 薊 馬雄成蟲在 三次 偏 好性選 擇實驗中 , 停 留 於健康 西瓜 (白色長 條 ) 及感染 WSMoV 之西瓜 (黑色長條) 植株上薊馬數量。(B) 薊馬雌成蟲在三次偏好性 選擇實驗中, 停留 於健康西 瓜 (白色長條) 及感染 WSMoV 之西瓜 (黑色 長條) 植株上薊馬數量。每次實驗重覆在兩植物間釋放 10 隻薊馬成蟲。在偏好 性實驗中 * 表示經符號檢定之 P < 0.05 以及 ** 表示 P < 0.01。誤差線 (error bars) 表示標準差 (standard error)。
Fig. 9. Feeding preference of Thrips palmi adults to thrips-damaged versus Watermelon silver mottle virus (WSMoV)-infected watermelon seedlings. (A) Number of male thrips adults on healthy (white columns) and WSMoV-infected seedlings (black columns) in three trials. (B) Number of female thrips adults on healthy (white columns) and WSMoV-infected seedlings (black columns) in three trials. Ten adults were released between the two plants at the beginning of the experiment. * and
** indicate significant differences (P < 0.05 and P < 0.01) between numbers of thrips adults on thrips-damaged and WSMoV-infected seedlings by sign test. Error bars indicate the standard error of the mean.
0 2 4 6 8
2 4 8 12 24
0 2 4 6 8
2 4 8 12 24
(A) (B)
** ** **
** **
** ** ** **
Number of thrips
Time after thrips release (hours) Time after thrips release (hours)
28 討論 1. 番茄斑萎病毒屬病毒的 傳播模式
南黃薊馬主要以持續性增殖型傳播模式傳播 WSMoV (表一、圖三)。一 齡幼蟲獲毒 後需等到 羽化為成蟲始得以傳播病毒,此類特殊的 stage effect 與西方花薊馬傳播 TSWV 相同。二齡幼蟲獲毒成蟲傳毒的比例大幅降低至 0.6%。Chen et al. (2006) 報導南黃薊馬可以以非持續性傳播方式 傳播,但本 研究結果顯示南黃薊馬亦可以以半持續性傳播,但無法以非 持續性傳播方式 傳播。在其它由西方花薊馬傳播 同為番茄斑萎病毒屬但不同血清群的病毒-
TSWV 及鳳仙花壞疽斑點 病毒 (Impatiens necrotic spot virus, INSV) 也有相 同的傳播模式與 stage effect。在西方花薊馬傳播 TSWV 及 INSV 例子中,
一及二齡幼蟲期皆可 獲得病毒,且於成蟲期傳播病毒,但是薊馬獲得病毒的 能力會隨薊馬分離株 不同而改變,例如 TSWV 之 NL3 分離株僅能由西方 花薊馬一齡幼蟲期獲毒且在成蟲期傳播病 毒,但 TSWV 之 IS2 與 US2 分 離株則可由一齡或二齡幼蟲期獲毒,並於 成蟲期傳播病毒 (van de Wetering et al., 1999a; Whitfield et al., 2005)。另外,煙草花薊馬的一齡與 二齡幼蟲也 可以傳播 TSWV (Sakimura, 1963)。東方番茄薊馬 (Ceratothripoides claratris) 傳 播 與 WSMoV 相 同 血 清 群 之 番 椒 黃 化 病 毒 (Capsicum chlorosis virus, CaCV),不管是一齡幼 蟲獲毒成蟲傳 毒、二齡幼蟲獲毒成蟲傳毒 及一齡幼蟲 獲毒二齡幼蟲傳毒, 均可有效地傳播 CaCV (Premachandra et al., 2005)。綜 合本研究中南黃薊馬 傳播 WSMoV 以及文獻中 TSWV、INSV及 CaCV 傳 播生態學研究,可以發現薊馬傳播番茄斑萎病毒屬的病毒一齡幼蟲獲毒比二 齡幼蟲獲毒於成蟲期的病毒傳播效率高,且隨薊馬分離株不同而有所差異。
南黃薊馬傳播 WSMoV 之傳播效率在性別間沒有顯著差異, 與東方番 茄薊馬傳播 CaCV 的情況相同 (Premachandra et al., 2005);但西方花薊馬 傳播 TSWV 時,雄蟲相較於雌蟲有較高的傳播病毒效率 (van de Wetering et al., 1999b; Stafford et al., 2011)。此外,TSWV 對西方花薊馬取食行為的影 響 亦有性別差異。Stafford et al. (2011) 比較健康雌薊馬與雄薊馬成蟲及感染 TSWV 雌薊馬成蟲與雄薊 馬成蟲的取食 電 位 (electrical penetration graphs),
29
結 果 發現感 染 TSWV 雄薊馬 取食頻率 高 於健康雄薊 馬,但 是感染 TSWV 雌薊馬取食頻率與健康雌薊馬則沒有差異。可能由於不同血清群 的病毒與不 同種類病媒薊馬的交互作用,所以 不同種類、不同性別的薊馬傳播病毒的效 率亦不盡相同。
2. 番茄斑萎病毒屬病毒對薊馬存活率與 發 育速率之影響
本研究結果指出 WSMoV 對南黃薊馬的存活率與發育速率沒有直接效 應。據 Whitfield et al. (2005) 指出過往的研究有兩大盲點:(1) 直接將健康 薊馬飼養於健康植物上,而將感染病毒的薊馬飼養於感染病毒的植物上,然 後比較存活率與發育速率等生活史特徵。由於感染病毒的植物及健康植物在 營養條件有所差異,這樣的比較會使 實驗結果受到植物營養條件的干擾。(2) 使用薊馬的非寄主植物做為薊馬取食的供試植物,如此薊馬在非寄主植物上 的適存度之影響將大於病毒效應,進而干擾實驗結果。DeAngelis et al. (1993) 比 較 飼養於感染 INSV 的植物上與飼養於健康植物 上的西方花薊馬 之存活 率與發育速率,結果發現取食染病植物對上述生活史特徵都有負面的影響。
此外,Robb (1999) 亦比較飼養於感染 TSWV 的植物上與飼養於健康植物 上之西方花薊馬發育速率之兩者的差異,結果發現取食染病植物 會延長西方 花薊馬的發育時間。 Wijkamp et al. (1996) 讓初生西方花薊馬幼蟲取食歐曼 陀羅 (Datura stramonium) 使其感染 TSWV,其後以西方花薊馬的非寄主植 物矮牽牛 (Petunia x hybrid ‘Blue Magic’) 葉片飼育,然後記錄薊馬幼蟲的 發育速率與存活率,並與未感染病毒之健康薊馬 幼蟲進行比較,結果發現兩 者的發育速率及存活率均無顯著差異。
除了 Whitfield et al. (2005) 點出以上兩大盲點外,我們亦發現,過往 針對植物病毒對病媒 薊馬的直接效應的相關研究,均使用可被病毒感染的寄 主植物,因而尚無法完全排除病毒與寄主植物之間交互作用的影響,所以精 確來說應當為間接效應。例如 Maris et al. (2004) 將健康西方花薊馬幼蟲以 健康及感染 TSWV 之甜椒葉片飼養,進行發育速率及存活率比較。結果發 現兩者之間存活率並 沒有顯著差異,但薊馬 幼蟲於感染 TSWV 之甜椒上的
30
發育速率較高。Belliure et al. (2005) 除了考量薊馬與寄主植物感染病毒與否 之 外 , 亦 考量 薊馬 取食 、機械傷害、薊馬接種 感染 TSWV、機械接種感染 TSWV 等 因子。 結果 發 現 健康 西方 花 薊馬 幼蟲 飼育 於 經薊馬取 食 過的 甜椒 比飼育於健康植物,其存活率較低 以及發育速率較慢;但西方花薊馬飼育於 薊 馬 接 種 以 及 機械 接 種 感 染 TSWV 的 甜 椒 則 與 飼 育 於 健康 甜 椒 者 存活 率 及發育速率沒有顯著差異。這顯示 TSWV 對西方花薊馬幼蟲有正向的效應,
但 是 此 效 應 主 要經 由影響 寄主 植物再 影響 取食於 其上的 西方 花薊馬 的存活 率與發育速率。病毒可能藉由抑制 薊馬叮咬 所引發的植物防禦機制,提供薊 馬較佳寄主植物品質。西方花薊馬協助 TSWV 傳播,TSWV 協助縮短病媒 薊馬幼蟲發育時間及 增加存活率,以促進西方花薊馬傳播 TSWV (Belliure et al., 2005)。阿拉伯芥 (Arabidopsis thaliana) 感染 TSWV 時,因薊馬 取食而 誘發的茉莉酸 (jasmonate, JA) 調控植物防禦機制會受到抑制,進而提高薊 馬幼蟲存活率 (Abe et al., 2012)。類似的現象也出現在 煙草粉蝨 (Bemisia tabaci) 傳播的番茄黃化捲葉病毒 (Tomato yellow leaf curl virus) 上 (Zhang et al., 2012),說明這樣的機制可能廣泛地出 現在持續性傳播模式 昆蟲媒介植 物病毒、病媒昆蟲與寄主植物的交互作用中。
雖然 Belliure et al. (2005) 的報告中區分清楚薊馬取食、機械傷害、薊 馬接種 TSWV、機械接種 TSWV 等各種因子,對薊馬幼蟲存活率與發育速 率之影響,可是在此報告中,並沒有逐一確認取食感染 TSWV 之甜椒的薊 馬 是 否 確 實感 染 TSWV,這樣會有因為薊馬幼蟲沒有感染病毒,而造成觀 察到的效應有所偏差的風險。在本研究中,WSMoV 對南黃薊馬幼蟲的發育 有間接效應 (圖六),當感染 WSMoV 的薊馬幼蟲以薊馬接種感染 WSMoV 的西瓜葉飼養時,其發育速率顯著高於以健康西瓜葉飼養者。值得注意的是 本 研 究 有 對 薊 馬 前 蛹 進 行 RT-PCR 前 測 試 , 確 認 實 驗 中 的 薊 馬 感 染 WSMoV 比例為 90%,所以我們可以排除因薊馬幼蟲未感染 WSMoV 而造 成 的 實 驗 誤差 , 此 外本 研 究 亦對 薊 馬 幼蟲獲 毒 來 源 西瓜 葉 片 進行 RT-PCR 檢測,確認直到薊馬幼蟲 AAP 結束時葉片都有可被偵測到的病毒濃度。
31
除了薊馬取食、寄主植物與機械傷害對薊馬 存活率與發育速率有所影響 外。病毒的分離株及病媒薊馬種類不同,也是重要影響因子。以感染 TSWV 之 CFL 分離株與 RG2 分離株 之紫背草 (Emilia sonchifolia) 及歐曼陀羅飼 養煙草花薊馬 ,結 果發現取食感染 TSWV 的植物令煙草花薊馬體型減小、
發育速率降低且存活率減低 (Stumpf and Kennedy, 2005)。以健康及分別感 染 TSWV 之 CFL 與 RG2 分離株的紫背草及歐曼陀羅為寄主植物飼養西方 花薊馬,結果發現當 西方花薊馬 取食感染 CFL 分離株之植物 與取食健康植 物相比,其發育速率提高、存活率增加;若西方花薊馬取食感染 RG2 分離 株之植物,雖然與取食健康植物相比只有發育速率提高,但存活率並沒有顯 著差異 (Stumpf and Kennedy, 2007)。本研究使用 WSMoV 的非寄主植物花 豆葉片飼養感染 WSMoV 及健康薊馬幼蟲,藉此完全排除病毒與寄主植物 間的交互作用 干擾,直接探討 WSMoV 與南黃薊馬之間的交互作用 。結果 發現 WSMoV 不管是對南黃薊馬的存活率或發育速率都沒有直接影響。
雖 然感 染 WSMoV 的南黃薊馬取食感染 WSMoV 的西瓜 葉片,其發 育速率 會提高 ,但是感染 WSMoV 的南黃薊馬於薊馬取食的西瓜葉片上發 育 速 率 最 高,以 及 在 健 康西 瓜 葉 片上發 育時間 最 長 (圖六 )。在西方花薊馬 與 TSWV 的案例中,西方花薊馬 於健康甜椒以及薊馬接種感染 TSWV 之 甜 椒 的 發育 速 率 均 較 於經 薊馬 取食 的 甜 椒 發 育 發 育速率 來得 高 。因 此有 關 WSMoV 與南黃薊馬幼蟲的關係仍需要進 一步實驗的澄清。
3. 南黃薊馬成蟲之取食偏好性
本研究分別使用不同性別南黃薊馬 成蟲進行偏好性實驗。在經薊馬取食 西瓜與健康西瓜偏好性實驗中及薊馬接種感染 WSMoV 的西瓜與健康西瓜 偏好性實驗中,顯示薊馬取食可以增加西瓜植株對雌、雄薊馬的誘引力 (圖 七)。另外,在薊馬取食與薊馬接種感染 WSMoV 西瓜偏好性實驗中,僅雄 薊馬對感染 WSMoV 的西瓜有較高的偏好性 (圖八);但雌薊馬則無偏好性,
研究的結果指 出 WSMoV 的感染對薊馬有吸引力且具性別上的差異 。類似 的 病 毒 感 染 所 產 生 的 性 別 差 異 影 響 , 也 出 現 在 西 方 花 薊 馬 及 其 所 傳 播 的
32
TSWV 上,TSWV 對西方花薊馬取食 行為 的影響有性別差異 (Stafford et al., 2011)。當雄薊馬感染 TSWV 時,其取食 頻率會增加,除了 取食頻率外,雄 薊馬發育時間亦會受到 TSWV 感染的影響。比較健康雄、雌薊馬之發育速 率,發現感染 TSWV 之雄薊馬發育速率高於健康者,可是雌薊馬沒有這種 現象 (Stumpf and Kennedy, 2007)。
雄 薊 馬 對 感 染 WSMoV 的 西 瓜 植 株 之 取 食 偏 好 性 可 能 源 於 植 物 對 抗 病原所產生的水楊酸 (salicylic acid),Abe et al. (2012) 發現當阿拉伯芥感染 TSWV 時 , 水楊酸 調控 植物 防 禦 機 制 相 關基因 之 表 現 會顯著 地 提 高 ,另外 將水楊酸塗佈於健康 阿拉伯芥葉面上時,會增加此植物對西方花薊馬的誘引 力。另外許多種類的雄薊馬在其腹部腹板 (abdominal sternites) 具有腹板腺 (sternal glands) 或縫孔區 (areae porosae),這些構造被認為與費 洛蒙的產生 有關係 (Mound, 2009)。花薊馬屬 (Frankliniella) 薊馬已有西方花薊馬、台 灣花薊馬 (F. intonsa) 和梳缺花薊馬 (F. schultzei) 三種花薊馬被報導會產 生聚集費洛蒙或性費洛蒙,吸引相同或不同性別薊馬聚集 (Milne et al., 2002;
de Kogel and van Deventer, 2003; Hamilton et al., 2005; Zhang et al., 2011)。南 黃薊馬形態上很可能也具有類似構造產生 的聚集費洛蒙或性費洛蒙,以吸引 雌薊馬進行交配。當雄薊馬被感染 WSMoV 的西瓜吸引而停留於植株上後,
雄薊馬可能會釋放與前述類似的費洛蒙以 吸引雌蟲前來交配,其後雌蟲會於 感染 WSMoV 的植株上產卵,讓薊馬幼蟲得以獲得病毒,待發育至成蟲期 時傳播 WSMoV。由此見得雄薊馬應於 WSMoV 流行病學中扮演不容忽視 的角色。
在本研究中,不論雄薊馬或雌薊馬對被南黃薊馬取食的西瓜,展現較高 的偏好性 (圖七)。這顯示薊馬取食會增加西瓜對薊馬的吸引力。據 Rhainds et al. (2007) 指出西方花薊馬取食菊花葉時 ,因為損害植物葉部 細胞膜,使 細胞內的類胡蘿蔔色 素 (carotenoid pigment) 外漏,使葉部呈現黃色。南黃 薊 馬 偏 好 經 薊 馬 取 食 過 植 物 , 可 能 是 由 於 薊 馬 對 黃 色 有 較 高 的 偏 好 性 (Mainali and Lim, 2011)。此外 El-Sayed et al. (2009) 指出紐西蘭花薊馬 (T.
obscuratus) 與 蔥 薊 馬 (T. tabaci) 對 植 物 受 薊馬 取 食 所 產 生 的 cis-jasmone