小蘗鹼對肺癌細胞生長抑制之研究; The investigation of berberine on cell growth inhibition of lung cancers
79
0
0
全文
(2) 謝 辭 年光似鳥翩翩過,想當初還懷著考取研究所的喜悅,如今,兩年的碩 士班時間已至尾聲。在中藥所中,我第一位認識也是在大學期間教導我實 驗方法,並鼓勵且幫助我繼續升學的闕副教授甫 ,即使現在她已遠赴美 國發展,依然不忘關心我的近況,於此,對於身處異鄉的啟蒙恩師獻上深 深的感謝與祝福。 初上研究所時,在闕老師的推薦下,得知台中榮總徐副教授士蘭博士 為一具專業知識及教學熱忱的老師,而徐老師亦願意指導我,因此,在這 兩年期間接受徐老師孜孜不倦的教誨,無論在學問的獲得或是處世的道理 上,均覺所獲良多。另外在所裡,謝校長明村博士也願意指導我,在謝校 長殷切的協助,並提供許多寶貴的意見下,方使本論文得以順利完成。在 此,對兩位指導教授致上學生最誠摯的謝忱與敬意。 此外,亦感謝吳教授午龍百忙之中評閱筆者論文,並願撥冗參與學生 口試,給予建言。而於研究期間承張永勳所長給予良好之學習環境,所內 陳忠川老師、謝文全老師、彭文煌老師、吳啟瑞老師以及其他所內老師, 均提供許多的知識及看法,對亦他們致上無限的謝意。 在實驗室中,非常感謝淑娥學姊、昀志學長、旗志學長、美君學姊、 淑鐘學姊、修竹學姊、張聰民學長、祖瑩學姊、郁婷學妹、亞雯學妹、佳 容、宜瑾、慶元與志學從旁的打氣和無私的協助,而同學兆群、炳賢、峻 銘、文雄、宏祺、冠宏和雯茹的鼓勵與幫助,亦送上最真誠的感謝。另外, 再謝謝每一位支持鼓勵我的親友。另外,要特別感謝這一路上陪伴著我, 一起相互扶持的素琴。 最後,僅將此論文獻給辛苦養育栽培我的父母親,沒有他們往日的用 心,怎有今日有小成的我。謝謝您們了!.
(3) 目錄 圖表目錄…………………………………………………………………… iii 中文摘要………………………………………………………………………I 英文摘要…………………………………………………………………… III 第一章 緒言………………………………………………………………… 1 第二章 總論………………………………………………………………… 4 第一節 小蘗鹼的文獻考察……………………………………………… 4 一、小蘗鹼之結構與理化性質…………………………………………4 二、小蘗鹼之藥用植物來源……………………………………………5 三、小蘗鹼之藥理學研究…………………………………………… 10 四、小蘗鹼之藥物動力學研究……………………………………… 16 第二節 細胞週期的調控…………………………………………………17 一、細胞週期………………………………………………………… 17 二、細胞週期調控分子……………………………………………… 18 第三節 MAPK 家族參與細胞週期之調控………………………………22 第四節 轉錄因子–NF-κB 與 AP-1…………………………………… 24 第三章 實驗材料與方法……………………………………………………27 第一節 實驗材料…………………………………………………………27 一、細胞株來源……………………………………………………… 27 二、實驗試劑………………………………………………………… 27 第二節 實驗方法與步驟…………………………………………………29 一、細胞培養………………………………………………………… 29 二、細胞數目的測定………………………………………………… 30 三、流式細胞分析儀測定…………………………………………… 30 四、蛋白質定性與定量的分析……………………………………… 31 i.
(4) 五、蛋白質激酶活性分析…………………………………………… 32 六、Gel Retardation Assay…………………………………………… 33 第四章 實驗結果……………………………………………………………36 一、觀察不同劑量小蘗鹼對肺癌細胞的影響……………………… 36 二、不同劑量小蘗鹼的生長抑制效應……………………………… 36 三、小蘗鹼使細胞週期停滯於 G1 期…………………………………37 四、小蘗鹼對細胞週期相關分子的調控…………………………… 37 五、小蘗鹼調控 MAPK 家族分子的活性……………………………38 六、小蘗鹼對蛋白質激酶活性的影響……………………………… 38 七、小蘗鹼對 AP-1 與 NF-κB 活性的影響………………………… 39 第五章 討論…………………………………………………………………40 第六章 結論…………………………………………………………………47 參考文獻…………………………………………………………………… 58. ii.
(5) 圖表目錄 圖 1. The structure of berberine. 4. 圖 2. The cell cycle control system. 21. 圖 3. The MAP kinases cascades. 24. 圖 4. Microscopic observation shows the anti-proliferation effect of berberine on four NSCLCs. 48. 圖 5. H1299, H1299/p53, A549 and CH27 cells were treated with drug-free medium(control), or medium containing 1, 10 or 100 µM of berberine for 12, 24, 48 and 72 hours. 49. 圖 6. Flow cytometric analysis of four NSCLCs treated with berberine(100 µM)for 24, 48 and 72 hours. 50. 圖 7. Western blot analysis of Cdks and cyclins. 51. 圖 8. Western blot analysis of CdkIs, p53 and retinoblastoma proteins. 52. 圖 9. Western blot analysis of MAPK family and phospho-Akt. 53. 圖 10. Kinase activitiy assay of Cdk2, Cdk4, Cdk6, ERK, JNK and p38 MAPK. 54. 圖 11. Gel shift assay of AP-1 and NF-κB. 55. 圖 12. Molecular mechanism of berberine on cell growth inhibition of NSCLCs. 56. 表 1. 含有小蘗鹼之藥用植物. 5. 表 2. four classes of Cyclins and Cdks in mammalian cell cycle. 19. 表 3. The effect of berberine on cell cycle distribution. 57. iii.
(6) 小蘗鹼對肺癌細胞生長抑制之研究 研究生. 梁峰賓. 中國醫藥學院 中國藥學研究所 藥理學組. 摘要 細 胞 週 期 調 控 系 統 中 , 最 重 要 的 分 子 為 cyclin-dependent kinases (Cdks),以及其活性調控蛋白 cyclins,藉由不同 cyclin-Cdk complexes 的組合可調節細胞進入不同的細胞週期,例如 cyclin D-Cdk4/6 以及 cyclin E-Cdk2 的活化可使得細胞週期由 G1 期進入至 S 期,而 cyclin B/Cdk 1 的活 化促進細胞由 G2 期進入 M 期,然而,經由與 Cdk inhibitors(CdkIs)結合, 這些活化型的 cyclin-Cdk complexes 的活性就被抑制,使得細胞週期的進 行停止。小蘗鹼(Berberine)為異奎啉(isoquinoline)類的植物生物鹼, 具有多方面的藥理作用,包含著抗發炎以及抗微生物的活性。過去的研究 已指出小蘗鹼於使用於血癌、胃癌與食道癌細胞上,具有抗腫瘤的效果。 儘管如此,但是關於小蘗鹼對人類非小細胞肺癌細胞(NSCLCs)的效應 未曾有文獻報導,且小蘗鹼抗腫瘤之分子作用機轉亦不清楚。於本實驗 中,以人類非小細胞肺癌細胞株 H1299 跟 H1299/p53 用來研究小蘗鹼藉由 調控細胞週期相關分子而使得細胞生長抑制的作用。以小蘗鹼(100 μM) 連續處理三天,肺癌細胞的生長明顯被抑制,且呈現劑量及處理時間依存 性的效果,並造成細胞週期停滯於 G1 期。由蛋白質西方墨點分析之結果 顯示小蘗鹼的生長抑制作用可能是與減少細胞內 cyclin D3、Cdk2 和 Cdk4 蛋白量的表現以及增加 cyclin E、p16 和 p21 的表現量有關。由於上述分子 在 蛋 白 質 表 現 量 上 的 變 化 使 得 Cdk2 和 Cdk4 激 酶 活 性 降 低 並 導 致 retinoblastoma protein-p130 無法被磷酸化。除此之外,ERK 蛋白量與活性 的減少可能亦牽涉其中。總而言之,小蘗鹼對於腫瘤細胞生長的抑制作. I.
(7) 用,是透過調控細胞週期相關分子 cyclins、Cdks 和 CdkIs 的表現量,而就 以上結果顯示,小蘗鹼可能為癌症治療提供另一新藥的來源。. II.
(8) The investigation of berberine on cell growth inhibition of lung cancers Fong-Pin Liang Institute of Chinese Pharmaceutical Sciences China Medical College. Abstract The central components of the cell-cycle regulatory system are cyclin-dependent kinases(Cdks), whose activity depends on association with regulatory proteins called cyclins. Oscillations in the activities of various cyclin-Cdk complexes leads to the initiation of various cell-cycle events. For instance﹐activation of cyclin D-Cdk4/6 and cyclin E-Cdk2 makes the cell-cycle progress from G1 into S phase.. However, the activities of cyclin-Cdk. complexes can be suppressed by the binding of Cdk inhibitor proteins(CdkIs). Then﹐the progression of cell-cycle may be paused. Berberine, an isoquinoline plant alkaloid﹐has multiple pharmacological actions﹐including anti-inflammatory and anti-microbial activities.. Although. berberine had been demonstrated to have antineoplastic function on leukemia﹐ gastric and esophageal cancer cells. Little is known about the cellular and molecular mechanisms of antitumor effect of berberine. In this study, human NSCLC cell lines H1299 and H1299/p53 were used to examine the antitumor effect of berberine. growth inhibition.. Treatment with berberine caused a dose-dependent Data from flow cytometry analysis indicated that. berberine-mediated antiproliferation effect related to G1 phase arrest. Moreover, this berberine-induced growth inhibition was associated with decrease the III.
(9) expression levels of cyclin D3、Cdk2 and Cdk4 proteins and increase in cyclin E、p16 and p21 protein.. In vitro kinase assay indicated that berberine. inhibited the Cdk2 and Cdk4 kinase activities and consequently lead to hypophosphorylated of retinoblastoma protein-p130. In addition﹐decrease in ERK protein level and activity is also observed. Taken together, berberine can inhibit cancer cell growth by directly modulating the expression of mutiple cell cycle regulatory molecules, such as cyclins、Cdks and CdkIs. These results suggest that berberine may be used as a potent agent for cancer therapy.. IV.
(10) 第一章 緒言 近幾年來,癌症的發生高居國人十大死因的第一位。世界衛生組織更 將癌症列為二十一世紀的三大殺手之一。最近的統計更顯示,在台灣約每 9 分 20 秒就發現一個新的癌症個案,其中肺癌(Lung Cancer)在男性人口 十大癌症發生率佔第二名,為女性人口死亡率的第一名,顯示到目前為 止,臨床上並無藥物可有效的控制及治療肺癌的發生。臨床上肺癌分為兩 大類,即小細胞癌(Small Cell Lung Cancers,SCLSs)與非小細胞癌 (Non-Small Cell Lung Cancers,NSCLCs) ,多數的患者屬於非小細胞癌, 肺非小細胞癌又可分為肺腺癌(adenocarcinoma lung cancer)、肺鱗狀細胞 癌(squamous cell lung cancer)和肺大細胞癌(large cell lung cancer),其 中肺腺癌為肺癌患者中最常見,無吸煙者所罹患肺癌常為此類,而肺鱗狀 細胞癌則常見於吸煙患者當中。 所謂的癌細胞,是因為體內細胞多次基因複製上的錯誤與突變而轉型 成的,於分子層次上是跳脫一般我們正常細胞受到的死亡與生長的調控機 制,癌細胞不易衰老、死亡且具有快速增殖的能力。在過去的研究發現, 許多刺激生長的接受體和訊息分子,如 erb-B、HER-2 和 Ki-ras、c-myc, 以及抗細胞凋亡的分子,像是 Bcl-XL,於癌細胞中均有大量表現或過度活 化的現象,這些分子我們稱之為致癌因子(oncogenes)。另一方面,一些 細胞週期中重要的調控分子,如 p53、p16 和 pRb,它們可能在基因庫上的 序列整段被移除,或是在 DNA 序列促進端(promotor)上被嚴重的甲基化, 也有可能是產生了突變,導致這些分子無法執行正常的功能,因而使細胞 週期漫無章法的進行著,促使癌細胞不斷的分裂增殖,所以,我們常稱這 些分子為腫瘤抑制因子(tumor suppressor genes)。當然,癌細胞的形成不 是只有單純的幾個分子失去調控而已,許多已知和未知的分子都有可能參 與其中,也由於癌細胞本身的複雜性,更增添了我們在癌症治療中的困難 1.
(11) 度。 日前,本校中國醫藥學院附設醫院曾利用中藥方劑三黃瀉心湯來治療 癌症病患,並宣稱有不錯的療效,而其中黃連(Coptis chinensis FRANCHET) 為三黃瀉心湯的主要組成藥材之一,黃連為中醫藥界廣為使用的植物性藥 材,其中小蘗鹼(berberine)為其主要的有效活性成分之一。小蘗鹼的發 現與研究已有數十年之久,在許多植物均有此化合物的存在,譬如台灣黃 蘗 ( Phellodendron wilsonii HAYATA ) 及 川 上 氏 小 蘗 ( Berberis kawakamii ,此外,其藥理活性亦是受到廣泛的探討與研究,包含有抗發炎、 HAYATA) 抗微生物、抗腦缺血以及抗心律失常等作用。至於在抗腫瘤活性方面,也 有不少人發表過這方面的研究,之前研究中多數均表明小蘗鹼會去影響到 癌細胞週期的變化,有的人認為是使細胞週期停滯於 G1 時期,也有人表 示是在 G2 期,另外,也有人提出小蘗鹼會誘導癌細胞的凋亡,然而也有 人說不會,形成了眾說紛紜的情形,當然,不同的癌細胞對藥物的感受性 有差異,也由於各種細胞間分子層次上表現的不一,而可能造成截然不同 的結果與現象是能夠預期的,因此,小蘗鹼對癌細胞的影響仍有很大的空 間去進一步探索與研究。 由於小蘗鹼對於非小細胞肺癌細胞所造成的影響仍不很清楚,於分子 層次上的調控機制也鮮少有人去了解與研究,因此,本實驗利用非小細胞 肺癌細胞株 H1299 與 H1299/p53 處理小蘗鹼,由於 H1299 肺癌細胞株本身 於基因庫上表達 p53 基因的序列已移除掉了,因此,便有人製造一個建構 有 p53 基因的質體送進去 H1299 細胞中,使之能表達正常功能的 p53 蛋 白,稱此肺癌細胞株為 H1299/p53, p53 蛋白是一眾所皆知的腫瘤抑制因 子,無論在細胞週期進行或是細胞凋亡過程中間,它均扮演著相當重要的 角色,便可利用在此分子上的差異性,比較小蘗鹼對兩細胞株是否造成不 一致的現象,使我們更能對小蘗鹼於癌細胞的作用與分子機轉有更深入的 了解,除此之外,也提供不管是癌症治療策略上或是抗癌新藥的開發上, 2.
(12) 另一個可能的途徑與方向。. 3.
(13) 第二章 總論 第一節 小蘗鹼的文獻考察 一、小蘗鹼之結構與理化性質:(1) 1. 別名:黃連素、Umbellatine。 2. 分類: 為 isoquinoline 類衍生物中 benzylisoquinoline 的 proberberine 型 的生物鹼。 3. 化學名: 5,6-dihydro-9,10-dimethoxybenzo[g]-1,3-benzodioxolo[5,6-a]quinol i-zinium 4. 結構式:. + 5. 分子式及分子量:[C20H18NO4] ;336.37. 6. 物理化學性質:. 小蘗鹼的羥基化合物為黃色針狀結晶,熔點 145℃,氯化物的二 水合物為黃色結晶。碘化物熔點 263∼265℃。UVMAX:225、270、 331nm。游離小蘗鹼能緩慢溶於水中,在冷乙醇中溶解度不大,但易 溶於熱水或熱乙醇,難溶於苯、氯仿和丙酮。 4.
(14) 二、小蘗鹼之藥用植物來源:(2–5) 表 1. 含有小蘗鹼之藥用植物 科名. 屬名. 植物名. 毛茛科 黃連屬 (Ranuncul 黃連 (Coptis) -aceae). 拉丁名 Coptis chinensis. FRANCHET. 所含部位 備註 根莖 (5.56~7. 25﹪). Coptis deltoidea C. Y. 三角葉黃連 根莖 CHENG et HSIAO. — 又名:峨 嵋連、峨 嵋家連. 雲南黃連. Coptis teetoides C. Y. CHENG. 根莖. —. 日本黃連. Coptis japonica MAKINO. 根莖 (6.7~9.2 ﹪). —. 五加葉黃連. Coptis quinquefolia MIQUEL. 根莖. —. Hydrastis canadensis LINNÉ. 根莖. —. 根. —. 北美黃連屬 (Hydrasti- 北美黃連 s). 唐松草屬 Thalictrum (Thalictru 箭頭唐松草 angustifolium LINNÉ m) 多葉唐松草. 又名:金 Thalictrum foliolosum 根及根莖 絲黃連 DE CANDOLLE. 高山唐松草. Thalictrum alpinum LINNÉ. 根. —. 狹序唐松草. Thalictrum atriplex FINET et GAGNEPAIN. 根莖. —. 偏翅唐松草. Thalictrum delavayi FRANCHET. 根. —. 大葉唐松草. Thalictrum faberi ULBRICH. 根. —. 尖葉唐松草. Thalictrum acutifolium 根莖 BOIVIN. —. 滇川唐松草. Thalictrum finetii BOIVIN. —. 5. 根莖.
(15) 香唐松草. Thalictrum foetidum LINNÉ. Thalictrum 金絲馬尾連 glandulosissimum FINET et GAGNEPAIN. 根. 根及根莖. Thalictrum glandulosissimum 昭通唐松草 FINET et GAGNEPAIN 根 chaotungense W. T. WANG et S. H. WANG 高原唐松草. Thalictrum cultratum WALLICH. 全草. — 又名:多 腺唐松草. 又名:昭 通馬尾連. 又名:草 黃連. 貝加爾唐松 Thalictrum baicalense 又名:球 根莖及莖 草 果白蓬草 TURCZANINOW 黃唐松草. Thalictrum flavum LINNÉ. 爪哇唐松草. Thalictrum javanicum 根及莖 BLUME. 根. — —. Thalictrum minus LINNÉ var. hypoleucum 東亞唐松草 莖及葉 (SIEBOLD et ZUCCARINI)MIQUEL. 又名:秋 唐松草、 小果白蓬 草. Thalictrum petaloideum LINNÉ. 根莖. 又名:腎 葉唐松草. 根. 又名:箭 頭唐松草. 瓣蕊唐松草. Thalictrum simplex 短梗箭頭唐 LINNÉ var. brevipes 松草 HARA 鞭柱唐松草. Thalictrum smithii BOIVIN. 小蘗科 小蘗屬 Berberis kawakamii (Berberida 川上氏小蘗 (Berberis) HAYATA ceae) 玉山小蘗 小蘗. 全草. 根及莖. — 又名:台 灣小蘗. Berberis morrisonensis 根及莖 — HAYATA Berberis thunbergii DE 根莖木質 又名:日 部 本小蘗 CANDOLLE. 6.
(16) 錐花小蘗. Berberis aggregata SCHNEIDER. 黃蘆木. Berberis amurensis RUPRECHT. 根及莖. 深黑小蘗. Berberis atrocarpa SCHNEIDER. 根. 直穗小蘗. Berberis dasystachya MAXIMOWICZ. 根(1.78 ﹪)及皮 又名:密 部(4.46 穗小蘗 ﹪). 鮮黃小蘗. Berberis diaphana MAXIMOWICZ. 又名:黃 根(1.72 刺、黃花 ﹪) 刺. 甘肅小蘗. Berberis kansuensis SCHNEIDER. 根(2.74 ﹪). —. 黑果小蘗. Berberis heteropoda SCHNEIDER. 莖皮(1.90 ﹪). —. 豪豬刺. Berberis julianae SCHNEIDER. 又名:老 根(3﹪)鼠刺、山 黃連. 細葉小蘗. Berberis poiretii SCHNEIDER. 根(1.16 又名:針 ﹪) 雀. 刺黑珠. Berberis sargentiana SCHNEIDER. 又名:黃 根(3.68 精刺、黑 ﹪) 石珠. 藍果小蘗. Berberis veitchii SCHNEIDER. 莖(1.08 ﹪). 貓刺小蘗. Berberis soulieana SCHNEIDER. 又名:假 根(2.31 蠔豬刺、 ﹪) 排骨筋. 匙葉小蘗. Berberis vernea SCHNEIDER. 根(1.58 ﹪). —. 粉葉小蘗. Berberis pruinosa FRANCHET. 莖(1.58 ﹪). —. 7. —. — 又名:大 葉小蘗 —. —.
(17) 廬山小蘗. 金花小蘗. 鮮黃連屬 (Jeffersoni 鮮黃連 a). Berberis virgetorum SCHNEIDER. 莖(2.84 ﹪)及根 又名:長 皮(4.72 葉小蘗 ﹪). Berberis wilsonae HEMSLEY. 又名:小 根(2.94 葉雞腳黃 ﹪) 連、川西 小蘗. Jeffersonia dubia (MAXIMOWICZ). 莖和葉. Mahonia japonica DE 根皮和葉 CANDOLLE. 翠峰十大功 Mahonia oiwakensis 勞 HAYATA. — —. 闊葉十大功 Mahonia bealei(FOR根和葉 勞 TUNE)CARRIERE. —. —. 無刺十大功 Mahonia shenii CHUN 莖 勞 南天竹屬 南天竹 (Nandina) 防己科 古山龍屬 (Menisper (Arcangeli 古山龍 maceae) sia). Nandina domestica THUNBERG. 薊罌粟. Arcangelisia gusanlung 莖及根 H. S. LO. Agremone mexicana. 8. 又名:甘 平十大功 勞 又名:沈 氏十大功 勞. 又名:闌 樹皮及根 天竹、南 天竺. 千金藤屬 Stephania cepharantha (Stephania 金線吊烏龜 塊根 HAYATA ) 薊罌粟屬. 又名:貓 兒頭、刀 瓜山樹、 角刺茶. 根和莖. 密葉十大功 Mahonia ganpinensis 勞 (LEVÉILLÉ)FEDDE. 罌粟科. —. BENCH. et HOOKER F.. 十大功勞屬 狹葉十大功 Mahonia fortunei (Mahonia 勞 FEDDE ) 十大功勞. 根及根莖. 全草. —. 又名:台 灣千金藤 —.
(18) LINNÉ. (Papaverac (Agremon eae) e) 白屈菜屬 (Chelidoni 白屈菜 um) 紫菫屬 (Corydalis 伏延胡索 ). Chelidonium majus LINNÉ Corydalis decumbens (THUNBERG)PER-. 地上部分. —. 塊莖. 又名:伏 生紫菫. Corydalis 對葉延胡索 ledebouriana KARELIN 塊莖 et KIRILOW. 又名:對 葉元胡、 薯根延胡 索. SOON. 蛇果黃菫. Corydalis ophiocarpa 全草 HOOKER F. et THOMSON. —. 齒瓣延胡索. Corydalis remota FIS塊莖 CHER et MAXIMOWICZ. —. 石生黃菫. Corydalis saxicola BUNTING. 全草. —. 延胡索. Corydalis yanhusuo W. 塊莖 T. WANG. —. 荷青花屬 (Hylomec 荷青花 on). Hylomecon japonica (THUNBERG)PRANTL 全草. 博落回屬 (Macleaya 博落回 ). Macleaya cordata (WILLDENOW)R.. 芸香科 黃檗屬 (Rutaceae (Phelloden 台灣黃蘗 ) dron). et KUNDIG. 又名:雞 蛋黃菜. 全草. —. BROWN 又名:魏 氏黃柏. Phellodendron wilsonii 樹皮 HAYATA et KANEHIRA. 黃蘗. Phellodendron amurense RUPRECHT. 樹皮(1.6 ﹪)及根 皮(9﹪). —. 黃皮樹. Phellodendron chinense SCHNEIDER. 樹皮. —. Taddalia asiatica (LINNÉ)LAMK. 根. 飛龍掌血屬 飛龍掌血 (Toddalia). 9. 又名:小 葉黃肉樹.
(19) 馬錢科 馬錢子屬 (Loganiac (Strychnos 呂宋果 eae) ). Strychnos ignatii BERG 種子. 鼠李科 棗屬 (Rhamnac 酸棗 (Ziziphus) eae). Ziziphus jujuba MILLER. 葉. 又名:海 南馬錢. —. 三、小蘗鹼之藥理學研究: 1. 抗腦缺血及中樞神經細胞保護作用 (1) 小蘗鹼可減少小鼠全腦缺血後海馬 CA1 區細胞凋亡的發生(6)。 (2) 小蘗鹼可拮抗 NMDA、H2O2 及無血清培養引起的大腦皮層神經 細胞損傷,其機制可能與減少膜脂質過氧化物生成,提高抗氧 化酶活性有關(7)。 (3) 小蘗鹼對 oxidative stress 損傷中樞神經細胞具有保護作用,其作 用機制可能與其提高抗氧化物質 GSH 含量,減少脂質過氧化物 有關(8)。 (4) 小蘗鹼對培養大鼠腦皮層元“缺血”性損傷具有保護作用,其 機制可能與小蘗鹼抑制“缺血”性損傷誘導的[Ca2+]i 異常升 高,減少脂質過氧化物生成,增加抗氧化物的含量有關(9)。 (5) 大鼠全腦缺血再灌注中,海馬 c-fos 呈現高表達。小蘗鹼的抗氧 化自由基及降低 c-fos mRNA 水平的作用可能是其抗腦缺血作 用機制之一(10)。 (6) 小蘗鹼致腦缺血兔血漿 TXB2 水平下降,是其抗腦缺血作用機 制之一(11)。 (7) 小蘗鹼可抑制谷胺酸誘導腦細胞 c-fos 基因表達過高及[Ca2+]i 的升高,為其治療腦缺血性的機制之一(12)。. 10.
(20) (8) 小蘗鹼對於去甲腎上腺素,高 K+及 H2O2 引起的[Ca2+]I 升高的 抑制作用,可能是其抗腦缺血作用機制之一(13)。 (9) 小蘗鹼可降低谷氨酸、去甲腎上腺素和 5-羥色胺引起的新生大 鼠腦細胞[Ca2+]i 升高,這可能是其抗腦缺血机理之一(14)。 2. 對毒蕈鹼型受體的作用 (1) 鹽酸小蘗鹼可活化毒蕈鹼受體,從而引起血管的內皮依賴性的 舒張反應及支氣管的收縮(15)。 (2) 小蘗鹼對大鼠唾液腺、大腦皮質和心臟的毒蕈鹼受体均有中度 親和力(16)。 3. 降血糖作用 (1) 對於高脂飲食造成大鼠明顯胰島素抵抗模型,小蘗鹼有與 Metformin 有相似的增加胰島素敏感性的作用(17)。 (2) 小蘗鹼能透過肝細胞發揮非胰島素依賴的降糖作用,但不能增 加胰島素的分泌(18)。 4. 抗血液凝集作用 (1) 小蘗鹼可抑制兔子血小板凝集作用,藉由抑制 arachidonic acid 的代謝,使 thromboxane A2 的合成下降(19)。 (2) 小蘗鹼可促進 plasminogen activator 引發的凝塊溶解(20)。 5. 血管舒張作用 (1) 於 離 體 兔 腸 動 脈 , 小 蘗 鹼 可 產 生 endothelium-independent relaxtion(21)。 (2) 低濃度的小蘗鹼(<10-6M)具有舒張離體兔主動脈作用(22)。 (3) 小蘗鹼能部分對抗 K+收縮豚鼠門靜脈平滑肌的作用(23)。 6. 心臟疾病改善的作用 (1) 小蘗鹼降低大鼠因 pressure-overload 所引發的心肥大(cardiac hypertrophy)症狀(24)。 11.
(21) (2) 小蘗鹼可使動作電位時間延長,對 IK1 及 IK 具有阻斷作用,亦可 顯著抑制 HERG channel(25)。 (3) 在豚鼠心肌細胞上,低濃度小蘗鹼即能抑制電壓依賴性鈣通 道,而高濃度時又能激動細胞內鈣庫釋放(26)。 (4) 小蘗鹼具有延長心肌再極化作用,可能藉由抑制 Iks 及增加 ICa 所造成的(27)。 (5) 在豚鼠心室肌細胞,小蘗鹼具有抑制 L 和 T 型鈣通道作用(28)。 (6) 小蘗鹼可能透過促進胞外 Ca2+內流和胞內 Ca2+釋放等途徑,增 高心肌細胞內游離 Ca2+濃度,顯示強心作用(29)。 (7) 小蘗鹼對於心肌缺血再灌注損傷的保護作用,是透過其維持 ATPase 的活性有關(30)。 (8) 於心肌細胞,小蘗鹼藉抑制 IK,增加 action potential duration(31)。 (9) 小蘗鹼有改善心肌缺血引發的心律不整作用,其作用可能是透 過抑制 KATP 通道的活化及縮短缺血期間動作電位與有效不反應 期時間(32)。 (10) 小蘗鹼能顯著縮小大鼠冠狀動脈結紮後 24 小時心肌梗塞範 圍,減少血清游離脂肪酸的增高,降低梗塞後病理性 Q 波的發 生率,顯示小蘗鹼對缺血心肌有保護作用(33)。 (11) 小蘗鹼對心肌 Ca2+內流有雙向調節作用,尤其在高 Ca2+條件 下,降低 Ca2+內流可能是其抗心律失常作用機制(34)。 7. 改善學習記憶作用 (1) 長期投與大鼠小蘗鹼,可改善 scopolamine 所致記憶障礙,其作 用可能是透過增加周邊及中樞 cholinergic neuronal system 的活 性(35)。 (2) 小蘗鹼 0.4、4μg/隻,側腦室注射可改善 scopolamine 所致記憶 獲得障礙及促進正常小鼠的記憶保持,對 cyclohexmide 致記憶 12.
(22) 再現障礙及開場行為則無影響(36)。 (3) 小蘗鹼對東莨菪鹼、戊巴比妥所造成的記憶獲得障礙有明顯的 改善作用,其作用可能與抑制腦內 AchE 有關(37)。 8. 對胃腸道的影響 (1) 小蘗鹼對結腸平滑肌細胞外鈣內流和細胞內鈣釋放均有抑制作 用(38)。 (2) 小蘗鹼可抑制人與大鼠結腸電子傳遞(ion transport)的作用(39) (40). 。. (3) 小蘗鹼可鬆弛大鼠胃底部的縱行肌(longitudinal muscle)(41)。 9. 免疫系統調節及抗發炎作用 (1) 小蘗鹼可誘導小鼠 macrophage interleukin-12 的產生,並且活化 p38 mitogen-activated protein kinase(MAPK),顯示小蘗鹼具有 免疫治療的活性(42)。 (2) 小蘗鹼對 BALB/c 小鼠實驗性自體免疫疾病 tubulointerstitial nephritis 有治療效果(43)。 (3) 通過抑制細胞表面黏附因子的表達而抑制多形核白細胞與內皮 細胞的黏附,可能是小蘗鹼發揮抗炎作用的機制(44)。 (4) 小蘗鹼可抑制小鼠脾細胞對 T-dependent mitogen convanavalin A 及 phytochemaggulutinin 的生長反應(45)。 (5) 小蘗鹼有抑制以二硝基氟苯所致小鼠遲發性超敏反應(DTH) 的作用,其機制可能是抑制了 IFN-γ、IL-1、TNF-α、IL-2 等細 胞因子的產生和分泌,從而抑制免疫反應,減輕炎症損傷(46)。 (6) 小蘗鹼的抗炎作用可能與抑制中性白血球趨化,產生活性氧的 功能,抑制自由基產生,降低 PLA2 活性,減少炎症組織中 PGE2 的產生等多因素有關(47)。 (7) 小蘗鹼可抑制 etoposide 及 camptothecin 誘發的胸腺細胞凋亡 13.
(23) (48). 。. (8) 小蘗鹼 10-4、10-5mol•L-1 可增強腹腔巨噬細胞和人全血白細胞 吞噬白葡萄球菌的功能;促進小鼠腹腔巨噬細胞產生 IL-1;抑 制脾細胞產生 IL-2 和抑制 T 和 B 淋巴細胞轉化;25、50mg• kg-1 加速小鼠網狀內皮系統對炭粒的廓清速率;抑制小鼠血清溶 血素的產生和足跖的 DTH 反應。顯示小蘗鹼可增強小鼠非特異 免疫反應,抑制細胞和体液免疫功能(49)。 (9) 小蘗鹼可抑制 phytochemaggulutinin、convanavalin A 及 pokeweed mitogen 促 lymphocytes 生長作用(50)。 10. 抗病原體作用 (1) 小蘗鹼有毒殺線蟲 Toxocara canis 幼蟲的效果(51)。 (2) 對 Staphylococcus aureus (ATCC 25993 and 6538P) 、Streptococcus sanguis(ATCC 10556)、Escherichia coli(ATCC 25922)和 Pseudomonas aeruginisa(ATCC 27853)小蘗鹼有抑制作用(52)。 (3) 對 一 些 口 腔 細 菌 Actinomyces naeslundii 、 Porphyromonas gingivalis 、 Prevotella intermedia 、 Prevotella nigrescens 及 Actinobacillus actinomycetemcomitans 等菌株有抑制生長作用 (53). 。. (4) 小蘗鹼具有抗瘧原蟲 Plasmodium falciparum 的活性(54)。 (5) 對 Staphylococcus aureus RN4220、S. aureus 50774、S. aureus P-20-CA 等菌株小蘗鹼均有抑制作用(55)。 (6) 小蘗鹼對 Helicobacter pylori 有生長抑制作用(56)。 (7) 對 Candida albicans、C. tropicalis 與 C. glabrata 等黴菌,小蘗鹼 有抑制作用(57)。 11. 抗腫瘤的作用 (1) 小蘗鹼可明顯的抑制 embryoid bodies 的 PECAM-1、MMP-1、 14.
(24) MMP-2、MMP-9 的表現,具有抑制腫瘤血管增生的潛力,另外 亦可降低細胞內 ROS 的量(58)。 (2) 對於打入 colon26/clone 20 adenocarcinoma 的小鼠,所引發的癌 症惡病質(cancer cachexia),小蘗鹼有改善作用,主要是抑制 IL-1 誘導的 IL-6 mRNA 的表現(59)。 (3) 小 蘗 鹼 併 用 抗 癌 藥 CPT-11 可 明 顯 抑 制 植 入 Lewis lung carcinoma(LLC)小鼠的腫瘤生長與淋巴轉移,抑制轉移機轉 為減少 activator protein-1(AP-1)的轉錄活性,使 urokinase-type plasminogen activator(u-PA)表現降低(60)。 (4) 小蘗鹼能抑制胃癌細胞 MGC-803 的生長,細胞週期停滯於 G0/G1 期,並誘發細胞凋亡(61)。 (5) 小蘗鹼可藉由抑制 cyclin B1 表現,導致 cdc2 的活性下降,使癌 細胞細胞週期停滯於 G2 期(62)。 (6) 對於人食道癌細胞株(human esophageal cancer cell lines:YES-1 to YES-6) ,小蘗鹼可使細胞週期停留在 G0/G1 期,而抑制細胞 的生長(63)。 (7) 在 小 鼠 及 大 鼠 身 上 投 與 致 癌 物 20-methylcholanthrene 或 N-nitrosodiethylamine(NDEA),小蘗鹼有保護的作用(64)。 (8) 小蘗鹼可抑制 YES-2 細胞 IL-6 的表現,顯示具有 anticachectic effect(65)。 (9) 小 蘗 鹼 可 抑 制 人 血 癌 細 胞 N-acetyltransferase. 活 性 及. HL-60. 的. Arylamine. 2-Aminofluorene-DNA. adduct. formation(66)。 (10) 小蘗鹼可抑制小鼠 Ehrlich ascites cancer cells 的 DNA 聚合酶活 性,此外對於人血癌細胞 HL-60 與人淋巴母細胞 CCRF-CEM 均有抑制作用(67)。 15.
(25) (11) 處 理 小 蘗 鹼 可 誘 發 人 血 癌 細 胞 HL-60 凋 亡 , 使 nucleophoamin/B23 mRNA 表現減少及端粒酶(telomerase)活 性下降(68)。 (12) 小蘗鹼可抑制人肝癌細胞 KIM-1 的 activator protein-1 活性。(69) (13) 對於 Balb/c 3T3 fibroblast cell,小蘗鹼可使細胞週期停滯於 G2/M 期(70)。 (14) 小蘗鹼對人血癌細胞 K562 有抑制生長的作用(71)。 (15) 小蘗鹼會誘發細胞週期停滯與細胞凋亡於人血癌細胞 HL-60 (72). 。. (16) 於人 teratocarcinoma cell NT2/D1,處理小蘗鹼會使 c-Ki-ras2 protooncogene 表現下降(73)。 12. 其他 (1) 小蘗鹼可抑制 MAO-A 的活性(74)。 (2) 小蘗鹼可鬆弛兔的 corpus cavernosum 及增加 intracavernous pressure(75)。. 四、小蘗鹼之藥物動力學研究: 1. 小蘗鹼與血清中白蛋白結合的動力學數值–在 25℃時,binding rate constant(k1)為 67.5(± 0.1) (mol liter-1)-1s-1;dissociation rate constant (k-1)為 1.7(± 0.1)× 10-3s-1;而 binding equilibrium constant 為 3.97 (± 0.06)× 104(mol liter-1)-1(76)。 2. 分析健康人口服鹽酸黃連素片,得體內藥物平均滯留時間(MRT) 為 32.63± 6.15 小時,累積藥尿排泄量(AUC)為 27.49± 4.74μg(77)。 3. 家兔口服投與小蘗鹼的動力學模式符合一室開放模式,其主要動力學 參數 T(peak)為 0.62± 0.25 小時,C(max)為 92.72± 50.89μg/L, 曲線下面積(AUC)值為 491.7± 295.5μg /L•h。結果表明,家兔口 16.
(26) 服小蘗鹼後能迅速吸收,且達到有效濃度(78)。 4. 健康人口服小蘗鹼(300mg/人)的動力學模式符合一室模式,T1/2(ka) 為 0.87± 0.026 小時,T1/2(ke)為 2.94± 0.13 小時,T(peak)為 2.37 ± 0.04 小時,AUC 值為 2799.0± 1128.5μg /L•h(79)。. 第二節 細胞週期的調控 一、細胞週期(Cell Cycle):(80) 一個細胞分裂增殖成兩個細胞,中間受到許多分子調控複製程序的進 行,這期間我們稱之為細胞週期,整個週期可被分為兩大時期–interphase (包含 G1、S、G2 phase)及 M phase,而在這過程當中,則會有三個 checkpoints 來決定細胞週期是否能進入至下一時期: G0 phase:又稱 senescence 或 quiescence,為細胞靜止期,經由某些分 子的刺激,可進行分化或進入 G1 期。 G1 phase:G 意指為 Gap,處於 M 期及 S 期的中間時期,細胞會複製 本身胞器與蛋白質,增大自體體積,準備進入 S 期,此期 間可長達數小時至數天之久。由 G1 要進入 S 期為第一個 checkpoint。 S phase:S 意指 Synthesis,此期細胞進行的是 DNA 的複製工作,需 10 至 12 小時的時間。 G2 phase:為 S 期與 mitosis 的中間時期,為進入 M 期作準備。而要由 S 進入到 M 期為第二個 checkpoint。 M phase:M 意指 Mitosis,為細胞週期最短的一期,進行細胞核與細 胞質的分裂,又可細分為數個時期,如下: Prophase:核內複製完的染色體進行濃縮,胞質中 mitotic 17.
(27) spindle 進行組裝,兩個 centrosome 分別移向兩 側。 Prometaphase:centrosome 移至 spindle pole 處,核膜破裂, 染 色 體 藉 由 kinetochores 附 著 到 spindle microtubules 上,並開始移動。 Metaphase:染色體排列於 spindle pole 中間的赤道板上,準 備進行分裂。此時為第三個 checkpoint。 Anaphase:複製完的染色體進行分裂成兩子染色體,此時 kinetochore microtubules 開始縮短,spindle pole 往外側移動。 Telophase:分裂完的子染色體移至 spindle pole 處,染色體 開 始 decondense , 新 的 核 膜 及 細 胞 質 中 contractile ring 開始形成 Cytokinesis:核膜形成完全,細胞質分裂為二,細胞準備重 新回到 interphase。. 二、細胞週期調控分子: 1. Cyclins and Cdks 調控細胞週期的進行主要為 Cyclins 與 Cdks( cyclin-dependent kinases)兩大家族,單獨的 Cdks 並無磷酸化其下游分子的能力,必須當 Cdks 與 Cyclins 結合形成複合體時,Cdks 才會具有蛋白質激酶的活性, 即可磷酸化下游分子;在細胞週期的四個時期中,都有特定促使每一時 期進行的 Cyclin-Cdk complexes,G1 期有 Cyclin D-Cdk4/6 complexes 以 及 Cyclin E-Cdk2 complexes,S 期有 Cyclin A-Cdk2 complexes,G2 期有 Cyclin B-Cdk1 complexes(81-83)。列表如下:. 18.
(28) 表 2. four classes of Cyclins and Cdks in mammalian cell cycle Cell-cycle stages Early/Mid G1 phase Late G1 phase S phase M phase. Cyclins Cyclin D1,D2,D3 Cyclin E Cyclin A Cyclin B. Cdks Cdk4﹐Cdk6 Cdk2 Cdk2 Cdk1(cdc2). 然而,就算 Cdks 與 Cyclins 結合在一起並不能代表一定就有活性, 當 Cdks 某些位置上被加上磷酸根時,有可能會使其失活性,例如在 Tyr15 的位置被 Wee1 kinase 磷酸化時,就會使 Cdks 失活性,相反的,在相同 位置 Cdc25 phosphatase 解磷酸化時,會使得 Cdks 又恢復活性;另外, 在 Thr14 如被 Myt1 加上磷酸根,同樣會使 Cdks 失活性,而在 Thr161 位置由 CAK(cyclin H/Cdk7 complex)加上磷酸根的話,可使得 Cdks 為活性型態,因此我們可以了解到,當 Cdks 上的 Thr14 和 Tyr15 有一者 或兩者均被加上磷酸根時,Cdks 活性是被抑制的,而 Thr161 單獨被磷 酸化,Cdks 才具活性(84)。 另一方面,Cyclins 的表現量是隨著細胞週期而有所變化的,在 G2 期 表 現 的 cyclin B 如 果 在 進 入 mitosis 的 晚 期 時 , 若 沒 有 被 anaphase-promoting complex(APC)ubiquitylation 分解,則會使 mitosis 停止於 anaphase;而在 G1/S transition 時的 cyclin E 如果大量表現,因為 要進入到 S 期需要 Cdk2 與 cyclin A 的結合,但大量的 cyclin E 把 Cdk2 都抓住了,因此便無法進入到 S 期(85)。 2. Cdk inhibitors and p53 Cdk inhibitors 顧名思義就是抑制 Cdk 活性的分子,CdkIs 主要可分 為兩大家族–CIP/KIP 家族與 INK4(means Inhibitors of Cdk4)家族, CIP/KIP 家族成員有 p21CIP1/WAF1/sdi1、p27KIP1 及 p57KIP2;INK 家族則有 p16INK4a、p15INK4b、p18INK4c 和 p19INK4d(86)。 一般而言,CIP/KIP 家族中的 p57KIP2 主要是跟細胞的分化比較相. 19.
(29) 關,而 p21CIP1/WAF1/sdi1 與 p27KIP1 對於所有的 cyclin-Cdk complexes 均有抑 制作用(87),它們會形成 cyclin-Cdk-CdkI 型式的三元體,藉由遮蔽掉 Cdks 的 catalytic site,使不能與下游分子反應,進而抑制了細胞週期的進行(88)。 另一方面,INK 家族則對於 G1 期的 Cdk4 與 Cdk6 具有選擇性的抑 制作用,它們的結構上含有 ankyrin repeat domains(89),會與 cyclin D 競 爭 Cdk4 跟 Cdk6 的結合區域,使 cyclin D 無法與 Cdk4 跟 Cdk6 形成有 活性的 complex,而使的細胞週期停滯於 G1 早至中期(90)。 p53 本身為一轉錄因子(transcription factor) ,平時與 MDM2 protein 結合在一起為不活化型,當細胞 DNA 受到傷害、缺氧(hypoxia)或是 致癌因子(oncogene)活化的形況下,就會誘使 p53 的活化,p53 參與 了包括細胞 DNA 的修復、細胞週期的停止與細胞凋亡(apoptosis)等的 生物反應,比如說 CdkIs 中的 p21CIP1/WAF1/sdi1 為 p53 主要下游調控分子, p53 的活化會使得 p21CIP1/WAF1/sdi1 於細胞內的表現量上升,導致細胞週期 的停滯,此外,有關細胞 DNA 修復的 GADD45 和 p53R2 以及細胞凋亡 相關的 Bax、NOXA、PUMA 和 p53AIP1 都會受到 p53 的調控(91)。 3. pRb and E2F families pRb(retinoblastoma protein, also called pocket protein)家族有 pRb (pRb1﹐p110)、pRb2/p130 與 p107 三個,E2F(early gene 2 factor)家 族則有 E2F-1 到 E2F-6(92), E2F 本身為 transcription factor,平時在與 Rb protein 結合的時候是沒有活性的,但當 Rb protein 被上游的 cyclin/Cdk complexes 磷酸化時,就會與 E2F 脫離,使 E2F 具活性。一般來說,Rb protein 本身有 16 個位置可能會被加上磷酸根,而 cyclin/Cdk complexes 主要會去磷酸化 Rb protein C-terminus 的位置,比如說 cyclin D/Cdk4 complex 會去磷酸化 Rb protein 在 Ser780、Ser795 與 Thr826 的位置(93)。. 20.
(30) 就目前所知,pRb 是跟 E2F-1,2,3 結合在一起,pRb2/p130 和 p107 是與 E2F-4,5 結合在一起,而 E2F-6 則和 pRb 較無相關,它主要與 Polycomb group proteins(PcG)結合在一起。一般認為,E2F-4,5 與 pRb2/p130 以 及 E2F-6 與 PcG 的結合是在 G0 與 early G1 期時,扮演 transcription repressors 的角色, E2F-1,2,3 則作用於 late G1 跟 S 期,為 transcription activators。近年來的研究發現,受到 E2Fs 家族調控的分子相當的多,除 了調控細胞週期進入 S 期的分子,如 cyclin E、cyclin A、c-Myb 和 Cdk2 外;尚有與 DNA 合成相關的,如 ORC protein、MCMs(mini chromosome maintenance proteins) 、Cdc6、ribonucleotide reductase、thymidine synthase 與 DNA polymerase α 以及與 DNA 修復相關的分子,如 msh2 和 mih1 (mismatch repair gene)﹐Fanconi anemia 和 rpa3(excision repair)與 rad51、recQ1 和 rad54(double-strand break recombination repair);另外 對於 mitosis 與 DNA-damage checkpoint、apoptosis 相關的分子,如 cyclin B、cdc2、Mad2、chk1、caspase-3 和 APAF-1,都會受到 E2Fs 家族的調 21.
(31) 控(94)。 因此,就最近的研究文獻報導,顯示 pRb 與 E2Fs 家族的分子,對 於細胞的影響不僅調控細胞週期,並且可能參與細胞分化與死亡的調控。. 第三節 MAPK 家族參與細胞週期之調控 Mitogen-activated protein kinase (MAPK)被認為是傳遞外界訊息至細 胞內的重要途徑,它們參與了細胞的生長、分化、轉型(transformation) 與凋亡(apoptosis)等生物現象(95)。於哺乳類動物細胞中,MAPK 家族可 分為三大類:extracellular signal-regulated kinase(ERK, also known as MAPK) 、C-Jun N-terminal kinse/ stress-activated protein kinase(JNK/SAPK) 和 p38 MAPK,它們均屬於 serine/threonine 的蛋白質激酶。另外,尚有一 蛋白質 ERK5(also known as BMK1, Big mitogen-activated protein kinase) 最近亦被認為是 MAPK 家族中的一員。通常 MAPK 活化的訊息傳遞包含 三個以上的蛋白質激酶參與,即 MAPK kinase kinase (MAPKKK), MAPK kinase (MAPKK) 和 MAP kinase (MAPK). (96). 。. 1. ERK ERK 包含兩個 isoforms 為 ERK1(p44 MAPK)與 ERK2(p42 MAPK) ,為 MAPK 家族中了解最多的。一般而言,Raf-MEK-ERK 路徑 的活化代表的是一細胞要進行生長、分裂和分化的訊號,當細胞膜上的 tyrosine kinase receptors 接受到生長因子(growth factors)或 mitogen 的 訊號時,便透過 Grb2 及 Sos 可去活化 p21ras,接著活化 MAPKKK (如 c-Raf-1),然後受到磷酸化的 MAPKK(如 MEK1 與 MEK2)會再磷酸 化下游的 p44 MAPK 和 p42 MAPK,被磷酸化的部位是 Thr-Glu-Tyr (TEY)motif,接著活化的 ERK 會移入細胞核中,接著會磷酸化其下 游的許多轉錄因子(transcription factor),例如 ternary complex factor 22.
(32) (TCF)Elk-1、serum response factor accessory protein Sap-1a、Ets1、 c-Myc、Tal 等。此外,一些細胞質中的蛋白質亦被認為是 ERK 的下游 分子,如 RSK (90KDa ribosomal S6 kinase, p90rsk, also known as MAPKAP-K1 )、 cytosolic phospholipase A2 和 microtubule-associated proteins(MAP),包含有 MAP-1、MAP-2、MAP-4 和 Tau(97)。 事實上,ERK 在細胞週期的調控也是佔有一席之地的,過去研究認 為,ERK 的活化並進入細胞核中,對於 cyclin D1 的轉錄是必須的(98)。 尚有文獻指出,Cdk2 移入核中(99)與其在 Thr-160 位置上的磷酸化(100) 亦受到 ERK 的影響。顯示 ERK 的活化是會促使細胞週期的進行。 2. JNK 當細胞受到外界的壓力或是一些 cytokine 的刺激時,便會去活化 JNK。JNK 主要有 3 種 isoform:JNK1、JNK2 及 JNK3,前兩者表現在 大部分的組織,而 JNK3 主要表現在腦部及睪丸(101)。JNK 主要被上游 的 MKK4(MAP kinase kinase 4)及 MKK7 所磷酸化,被磷酸化的部位 是 Thr-Pro-Tyr(TPY)motif。活化的 JNK 可與 c-Jun 的 NH2-termianl activation domain 結合,並在 c-Jun 的 Ser-63 和 Ser-73 位置接上磷酸根, 接著活化的 c-Jun 可增加 AP-1 相關基因的表現。目前認為 JNK 下游的 分子包含 c-Jun、ATF-2 (activating transcription factor 2) 、 Elk-1、p53、 DPC4、Sap-1a 和 NFAT4(102)。 3. p38 MAPK 當 細 胞 受 到 紫 外 線 、 lipopolysaccharide 以 及 pro-inflammatory cytokines(如 IL-1 和 TNF-α)影響時,p38 MAPK 便會活化。p38 MAPK 主要有 4 種 isoform:p38α、p38β、p38γ 以及 p38δ,它們均會受到 MAPK kinase MKK6 (SKK3)和 MKK3 的磷酸化,被磷酸化的部位是在 Thr-Gly-Tyr(TGY)motif。p38 MAPK 在生長、分化、凋亡和 cytokine 生成的調控上,佔有很重要的角色。活化的 p38 MAPK 可去磷酸下游的 23.
(33) 轉錄因子(transcription factors) ,如 Stat1、ATF-2、Sap-1a 和 GADD153 (growth arrest and DNA damage transcription factor 153)(97)。 在 p38 MAPK 與細胞週期的相關性上,一般認為,p38 MAPK 扮演 一個負向調節的角色,它可抑制 cyclin D1 的轉錄作用(98)。另有研究指 出,p38 MAPK 可藉由影響 Cdc25 phosphatases 的活性,使細胞週期停 滯於 G2 期(103)。. 第四節 轉錄因子–NF-κB 與 AP-1 一、NF-κB Nuclear factor-κB(NF-κB)為一轉錄因子(transcription factor) ,表現 於 大 多 數 的 細 胞 當 中 。 結 構 上 , NF-κB 於 amino-terminal 區 域 含 有 24.
(34) Rel-homology domain(RHD) ,此區域中包含 DNA-binding 和 dimerization domains,以及含有 nuclear localization sequence(NLS)。到目前為止,至 少有 5 種 NF-κB 家族分子於哺乳類細胞中發現,分別是 RelA(also known as p65) 、c-Rel、RelB、NF-κB1(p50/p105)與 NF-κB2(p52/p100) ,其中 NF-κB1 和 NF-κB2 是分別由 105 kDa 與 110 kDa 的大分子前趨物 (precusors)轉變而來的具活性小分子,而它們兩者本身可自行形成 homodimers 進行調控,其餘的 NF-κB 家族則需形成 heterodimers 才具有作 用。 NF-κB 位於細胞質時,是與一抑制性分子結合成不活化型,稱為 IκBs, IκBs 家族主要有 IκBα、IκBβ 以及 IκBε,它們結構上含有 ankyrin-like repeat domains,可遮蔽掉 NF-κB 的 NLS 區域,使 NF-κB 無法進入細胞核中進行 調控。當一些刺激來臨時,如 pro-inflammatory cytokines(TNF-α 或 IL-1 β) ,可誘發一連串的訊息傳遞,使得兩個主要的 IκB kinases 活化,為 IKK-1 (IKKα)以及 IKK-2(IKKβ),它們會磷酸化 IκBs 在特定 amino-terminal serine residues(Ser32 和 Ser36 在 IκBα;Ser19 和 Ser23 在 IκBβ),被磷酸 化的 IκBs 會被 E3 ubiquitin ligase 所 ubiquitination,接著 IκBs 就會被 26S proteasome 所分解掉,如此一來,NF-κB 便成為自由態,即可進入細胞核 中與特定的 DNA 序列結合,以進行調控(104)。. 二、AP-1 Activator protein-1(AP-1)是最早被定義出來為轉錄因子(transcription factor)的其中之一,它調控了許多細胞的生物反應,包括生長、死亡,存 活和分化。AP-1 並不是一單獨的蛋白分子,而是由一群 basic region-leucine zipper(bZIP) proteins 所形成的二聚體(dimer)分子,這些分子有 Jun (c-Jun、JunB 和 JunD) 、Fos(c-Fos、FosB、Fra-1 和 Fra-2) 、Maf(c-Maf、 MafB、MafA、MafG/F/K 和 Nrl)以及 ATF(ATF-2、LRF1/ATF3、B-ATF、 25.
(35) JDP1 和 JDP2)四類家族,一般認為,它們所認知的區域為 12-O-tetradecanoylphorbol-13-acetate(TPA)response element(5’-TGAG/CVCA-3’) 或是 cAMP response element(CRE,5’-TGACGTCA-3’)。目前已知,Jun 與 Fos 這兩大家族是最為廣泛探討及研究,其中 Fos 家族不能自己家族內 形成 homodimers,必須要和其他家族形成 heterodimers,而 Jun 家族則無 此限制,可形成 homodimers 或 heterodimers(105)。. 26.
(36) 第三章 實驗材料與方法 第一節 實驗材料 一、細胞株來源: 本文所使用之細胞株 H1299、H1299/p53、A549 和 CH27 為人類非小 細胞肺癌細胞株(Human Non Small Cell Lung Cancer),由美國組織細胞 庫(ATCC)購得。其中 H1299 與 H1299/p53 屬於肺大細胞癌細胞株(large cell lung cancer cell lines) ,A549 為肺腺癌細胞株(lung adenocarcinoma cell line) ,而 CH27 則為肺鱗狀細胞癌細胞株(squamous lung cancer cell line) 。. 二、實驗試劑: 1. 榮民製藥: Penicillin﹐Streptomycin 2. ABI(American Biorganics INC): Tris 3. Amersham: ECL detection kits 4. BD PharMingen: Anti-cyclin A monoclonal antibody﹐Anti-cyclin E monoclonal antibody 5. BD Transduction Laboratory: Anti-Cdk1 monoclonal antibody ﹐ Anti-Cdk2 monoclonal antibody ﹐ Anti-Cdk4 monoclonal antibody﹐Anti-cyclin B monoclonal antibody﹐ Anti-cyclin D1 monoclonal antibody ﹐ Anti-cyclin D3 monoclonal antibody﹐Anti-pRb1 monoclonal antibody﹐Anti-pRb2/p130 monoclonal antibody 6. Bio-Rad: 27.
(37) Acrylamide﹐Ammonium Persulfate(APS)﹐ Glycine﹐Kaleidoscope Prestained Standards ﹐ N,N’-methylene- bis-acrylamide ( Bis )﹐ N,N,N’,N’,-Tetra-methylethylenediamine(TEMED) 7. CALBIOCHEM: Anti-Erk1/2 polyclonal antibody﹐Anti-Erk1/2 phospho-specific polyclonal antibody ﹐ Anti-p38 polyclonal antibody ﹐ Anti-p38 phospho-specific polyclonal. antibody ﹐ Anti-SAPK/JNK. polyclonal. antibody ﹐. Anti-SAPK/JNK phospho-specific polyclonal antibody 8. Falcon: Cell culture dish (10cm diameter)﹐Cell culture plate (12 well) 9. Gibco: RPMI 1640 Medium 10. Hyclone: Fetal Bovine Serum (FBS) 11. NEN: L-glutamine ﹐ Non Essential Amino Acid Solution (NEAA) ﹐ Polyvinylidene fluoride(PVDF) Transfer Membrane﹐Trypsin 12. Merck: 2-Mercaptoethanol ( 2-ME )﹐ EDTA ( C10H14N2Na2O8 . 2H2O )﹐ Dimethylsulfoxice (DMSO)﹐Methanol﹐Sodium dodecyl sulfate (SDS)﹐ Sodium chloride (NaCl)﹐Xylene cyanol 13. Pharmacia: Protein A sepharose 14. Santa Cruz Biotechnology﹐Inc: Anti-CDK6 polyclonal antibody﹐Anti-cyclin D2 monoclonal antibody﹐ Anti-p15 polyclonal antibody﹐Anti-p16 polyclonal antibody﹐Anti-p27 28.
(38) polyclonal antibody 15. Sigma: Berberine hemisulfate﹐Dithiothreitol (DTT)﹐Leupeptin﹐Methylene blue (C16H18ClN3S.3H2O)﹐Propidium Iodide(PI)﹐Sodium Pyruvate 16. Upstate Biotechnology: Anti-Akt phospho-specific monoclonal antibody﹐Anti-p21 monoclonal antibody﹐Anti-p53 monoclonal antibody. 第二節 實驗方法與步驟 一、細胞培養: 1. 細胞培養液的配製 將 RPMI 1640 粉末先溶於約 9.5 公升 Mili-Q 去離子水中,加入 20 g 的 NaHCO3,再用 HCl 調整 pH 至 7.0~7.2 後以 Mili-Q 去離子水補足體 積至 10 公升,然後以 0.22 µM 的過濾器過濾滅菌,最後儲存於 4˚C 冰 箱備用。 2. 完全培養液 每 500 毫 升 RPMI 1640 細 胞 培 養 液 添 加 100 IU 青 黴 素 ( Penicillin )、 100 μ g 鏈 黴 素 ( Streptomycin )、 2 mM 麩 胺 酸 (L-glutamine) 、100 μM 非必須胺基酸 (non-essential amino acid), 1mM 焦葡萄酸鈉(Sodium Pyruvate),最後加入 10%胎牛血清(Fetal bovine serum,FBS)即得。 3. 細胞株的培養 H1299、H1299/p53、A549 和 CH27 四種非小細胞肺癌細胞株,均 以 RPMI 1640 完全培養液培養於 37℃、5%CO2 的培養箱中;繼代培 養(subculture)時,先用 PBS(Phosphate-Buffered Saline)清洗 1 至 29.
(39) 2 次,再以 TEG (0.05 mM trypsin、2.5 mM EDTA 和 2.8 mM glucose) 處理數分鐘,使細胞與培養皿底解離,最後分種於新的細胞培養皿中, 每一培養皿注入約 7 毫升的細胞培養液,細胞培養液每兩天更換一次。. 二、細胞數目的測定: 將培養於細胞培養皿的細胞經 TEG 處理過後,以 3~5 毫升的培養液 將細胞打下來,使用血球計數器(hemocytometer)計算所有的細胞數目, 接著以每 ml 有 3x104 個細胞分種於 12 孔細胞培養盤中,每孔加入 1 毫升 細胞與培養液的混和液,均勻搖散細胞後,將細胞培養於 37℃、5%CO2 的培養箱中,經 24 小時後給予各濃度藥物(1μM﹐10μM﹐100μM) 處理,再經 12、24、48、72 小時後,以血球計數器計算各孔盤內的細胞 數,每一藥物濃度處理為三重覆。. 三、流式細胞分析儀測定:(62) 將細胞以每 10 毫升有 4x105 個細胞種於 10 公分的培養皿中,在藥物 分別處理 24、48、72 小時後,先以 PBS 清洗細胞,再將細胞用 TEG 處 理,用 1 毫升 PBS 細胞打下來,置於 1.5 毫升的 eppendorf tube 中,再以 900 rpm 室溫下離心 5 分鐘,倒掉上清液,置於震盪器輕輕震盪,將 1 ml 的 70% EtOH/PBS 一滴一滴滴入 eppendorf tube 中,使細胞完全均勻分散 於 70% EtOH/PBS 中,即可置於 4˚C 冰箱保存。 待要分析前,將 sample 以 900 rpm 在 4℃下離心 5 分鐘,倒掉上清液, 於避光條件下,每一個 sample 重新分佈於 1 毫升 PI solution 中,置於 37 ℃中作用 30 分鐘,最後置於冰上,利用 Flow cytometer(FACScan)儀器 分析,所得之結果以 ModFIT LT 2.0 軟體分析,加以量化之。. 30.
(40) 四、蛋白質定性與定量的分析: 1. 蛋白質的萃取(Protein Extraction) 將細胞用冰的 PBS 洗二次後用刮棒刮下,於 4℃下 5000 rpm 離心 5 分鐘,倒掉上清液並用棉棒將多餘的水吸乾,加入適量的 RIPA buffer 〔50 mM Tris(pH 7.4)﹐150 mM NaCl﹐1 ﹪NP-40﹐0.25 ﹪Na Deoxycholate﹐5 mM EDTA (pH 8.0) ﹐1 mM EGTA (pH 8.0) ﹐* 1 mM DTT﹐*5 µg/ml Leupeptin﹐* 0.2 mM PMSF﹐* 5 µg/ml Aprotinin﹐* 1 mM Na Vanadate﹐* 1 mM NaF(* add before use)〕 ,將細胞 pellet 均勻打散, 置於冰上作用 20 分鐘,接著以超高速離心 55000 rpm、4℃、30 分鐘, 收集上清液,以 Bradford 的方法測量蛋白質溶液,在波長 595 nm 下 的吸光值,換算成蛋白質濃度(μg/µl) 〔以一系列已知濃度的 BSA 做 成之 stand curve 換算蛋白質濃度。以每個 eppendorf tube 50µg 的蛋白 質分裝並用 RIPA buffer 調整成相同體積,接著再加入調整完體積的蛋 白質溶液 1/3 量的 4X protein loading dye(8 % SDS﹐0.04 % Serva blue R-250﹐40 % glycerol﹐200 mM Tris pH 6.8﹐10 % 2-mercaptoethanol ) , 以 95℃乾浴加熱變性 10 分鐘後,即可置於-80℃中保存。 2. 聚丙烯醯胺膠體電泳法(SDS-PAGE Assay) 將蛋白質依分子量分離利用 SDS-PAGE,首先配製 1.5 mm 厚的 discontinuous acrylamide gel,gel 分上下兩層,下層 separating gel 其 acrylamide 的百分比視分析蛋白質分子量而定,上層的 stacking gel 則含 4 % acrylamide。配製完成的膠體放置於電泳槽內,加入電泳緩衝 液(running buffer:25 mM Tris﹐192 mM glycine﹐0.1% SDS) 。接著 將萃取出的蛋白質與 4X protein loading dye 混合液再以 95℃乾浴 加 熱變性 10 分鐘,冰浴冷卻後離心。將各 sample 及標示標準分子量的 Multimaker 依序注入膠體的孔槽中,通以電壓 80 Volt.,待樣品通過 31.
(41) stacking gel 後電壓調整為 100 Volt.,當 serva blue R-250 的 dye 跑到 膠片底部或視其需要,即可將電源關閉。 3. 西方轉漬法(Western blot )(90) 將 PVDF membrane 浸於 methanol 數秒後以 Milli-Q 水浸濕,接著 裁好的濾紙與 membrane 先浸泡在 transfer buffer (50 mM Tris﹐40 mM glycine﹐0.375 % SDS﹐pH 9.0-9.4;20 % methanol )中。裁下電泳膠 中所要轉漬的區域後,亦浸泡於 transfer buffer 中。轉印石墨板之正極 板以 transfer buffer 潤濕後,依序重疊平鋪 4 張濾紙、membrane、gel、 4 張濾紙,最後蓋上以 transfer buffer 潤濕的負極石磨板,設定電流(電 流=濾紙面積 × 9/7 × 疊數),通電流經 1 小時又 10 分鐘後將 membrane 取出,浸泡於 5 ﹪non-fat milk/TBST 中,於室溫下搖晃至 少 30 分鐘。以 TBST buffer(24.22 g Tris﹐87.75 g NaCl﹐10 ml Tween 20﹐加水調到 1L )清洗 membrane 10 分鐘三次,加入一級抗體,並 置於 4℃下作用 overnight。 隔日先以 TBST buffer 清洗 membrane 10 分鐘三次,加入二級抗 體,使其在室溫下搖晃作用 1 小時之後,再用 TBST buffer 清洗 10 分 鐘 三 次 , 接 著 在 暗 房 中 將 membrane 與 ECL ( enhanced chemi-luminescence)反應後,裝於透明塑膠袋內並置於壓片夾中,以 X-ray film 感光顯影,再以自動沖片機沖片。. 五、蛋白質激酶活性分析(Kinase Activity Assay)(99) 將細胞用冰的 PBS 洗二次後用刮棒刮下,於 4℃下 5000 rpm 離心 5 分鐘,倒掉上清液並用棉棒將多餘的水吸乾,加入適量的 lysis buffer (RIPA buffer:Tris buffer=1:1﹐*200 µM PMSF﹐*20 µl/ml Leupeptin﹐ *1 mM NaF﹐*1 mM Sodium Vanadate) (*add before used) ,依照上述萃取 32.
(42) 蛋白質的步驟,取定量蛋白質(250-350μg)以 Tris buffer 調整體積到 250 或 300 μl。蛋白質溶液加入 4 μl 的 antibody 以及 30 μl protein A sepharose 後,在 4℃冰箱內旋轉作用 over night。 次日混合液以 7000 rpm 4℃離心 10 分鐘,吸掉上清液後,再加入 400 μl buffer Ι 〔20 mM Tris (pH 7.4) ﹐0.5 M NaCl〕 清洗 3 次,接著 buffer ΙΙ 〔20 mM Tris (pH 7.4)﹐0.5M DTT﹐20 µg/ml Leupeptin, 20μM PMSF﹐ 1 mM Sodium Vanadate﹐1 mM NaF〕 清洗 1 次,每次清洗於 4℃下以 7000 rpm 離心 5 分鐘,完成後吸掉上清液。 接著每管 sample 加入 15μl reaction mixture 〔50 mM Tris (pH 7.4)﹐ 10 mM MgCl2﹐0.5mM DTT﹐5μM ATP﹐1μg /μl Histone H1 、 RB protein、MBP or c-Jun ﹐2μCi γ-32p ATP〕 在 37℃作用 20 分鐘之後, 加入 4X protein loading dye 在 95℃下加熱變性 15 分鐘,將 sample 離心 後,注入 12.5 % SDS-PAGE gel 進行蛋白質電泳,待 Serva blue R-250 的 dye 跑到膠片底部,即可將電源關閉。依照西方轉漬法的步驟,將蛋白質 轉印到 PVDF membrane 後,將 membrane 裝於透明塑膠袋內並置於片夾 中,在暗房中以 X-ray film 直接感光顯影,再以自動沖片機沖片。. 六、Gel Retardation Assay(Gel Shift Assay):(108) 1. 細胞核的萃取 將細胞用冰的 PBS 洗二次後刮下,在 4℃下 10000 rpm 離心 5 分 鐘,倒掉上清液並用棉棒將多餘的水吸乾,加入適當的 Buffer 1 ( 50 mMTris-HCl/pH7.5﹐4 mM EDTA/pH8.0﹐2 mM EGTA ﹐ddH2O) 混 合均勻,置於冰上作用 20 分鐘後,將細胞混和液以 Trypan blue (0.4W/V)染色,看細胞膜是否有破裂,鏡檢下當 Trypan blue 可染入 細胞質中,即表示細胞膜已有破裂,接著在 4℃下 10000 rpm 離心 20. 33.
(43) 分鐘,倒掉上清液,再加入適量 Tortex Buffer(ddH2O﹐20 mM Hepes/pH7.9﹐350 mM NaCl﹐20% Glycerol﹐1% NP-40﹐5 mM MgCl2﹐ 0.5 mM EDTA/pH8.0﹐0.1 mM EGTA﹐*0.5 mM DTT﹐*0.1% PMSF﹐ *1% Aprotinin) (*add before use) 混合均勻,置於 4℃冰箱旋轉 over night。 隔日在 4℃下 13000 rpm 離心 15 分鐘,收集上清液,以 Bradford 的方法測量蛋白質溶液,在波長 595 nm 下的吸光值,換算成蛋白質 濃度(μg/µL),以每個 eppendorf tube 10 µg 的蛋白質分裝,置於-80 ℃保存。 2. Biotin probe 的配置 將互補的 DNA (biotin-AP-1 or biotin-NFκB-1) 單股加在一起, 以 ddH2O 配置成 5 pmole 的濃度,在 37℃下作用 1 小時,然後置於室 溫下 2 小時即得。 3. 反應與跑電泳 先將 6%的 native acrylamide gel 〔ddH2O 7.33 mL, 5x TBE buffer 2.5 ml, 30%Acrylamide/bis (29:1)2.52 ml﹐10%APS 125μl﹐TEMED 25μl〕 ,以 1x TBE buffer 作為 running buffer,在 12℃冷循環分析條件 下,用每片 gel 20 mA 預跑(pre-run)1 個小時。接著將萃取出的核蛋 白,每個 sample 加入 2μl 的 buffer D (ddH2O﹐20 mM Hepes/pH7.9﹐ 20% glycerol﹐100 mM KCl﹐0.5 mM EDTA/pH8.0﹐20% NP-40﹐*2 mM DTT﹐*0.1% PMSF ) (*add before use)及 4μl 的 buffer F(ddH2O﹐ 20% Ficoll 400﹐100 mM Hepes/pH7.9﹐300 mM KCl﹐*10 mM DTT﹐ *0.1% PMSF) (*add before use)以及 5μl 的 biotin-probe,然後在 30℃下作用 30 分鐘,完後加入作用完 sample 五分之一體積的 6x sample buffer (ddH2O 0.3 ml, 100% glycerol 0.5 ml﹐5x TBE Buffer 0.2 ml﹐ Xylene cyanol),接著將 sample 注入膠體的孔槽中,在 12℃下以每片 34.
(44) gel 30 mA 跑膠,待大約 2 小時後,6x sample buffer 跑到膠體的最底端 即可。下一步便利用上述西方轉漬的方法,以 0.5x TBE buffer 作為 transfer buffer,150 mA 下 1 個小時 transfer 到 NC membrane 上,然後 將 membrane 用 UV light 照射,接下來用 5% non-fat milk/1x TBS 室 溫下 blocking 30 分鐘,再用 1x TBS 清洗 membrane 5 分鐘 4 次,接著 將 membrane 浸泡於 1:20000 的 0.5μg/μl Streptavin-Horseradish Peroxidase conjugate in 1x TBS,在 4℃下 overnight。 次日,將 membrane 以 1x TBS 清洗 5 分鐘 4 次,接著在室溫下在 membrane 上加入 SuperSignal Substrate 作用 5 分鐘,然後將 membrane 裝於透明塑膠袋內並置於片夾中,在暗房中以 X-ray film 直接感光顯 影,再以自動沖片機沖片。. 35.
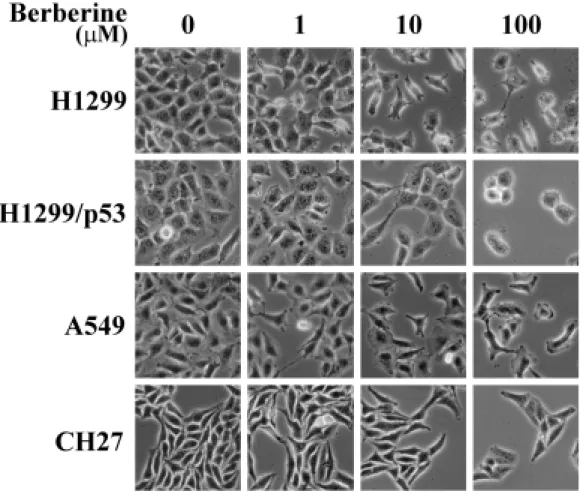
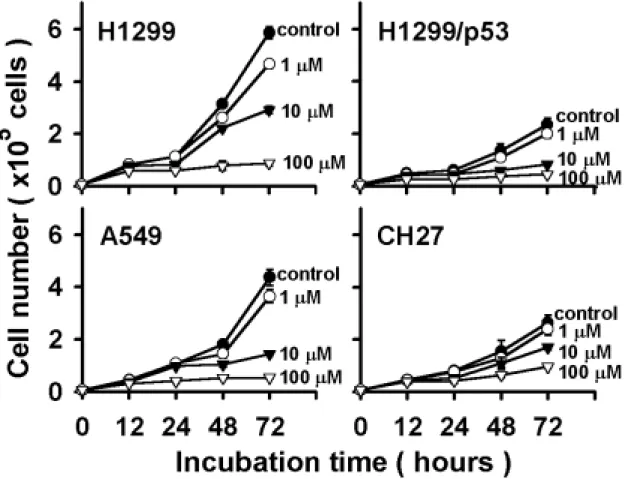
(45) 第四章 實驗結果 一、觀察不同劑量小蘗鹼對肺癌細胞的影響: 為了探討小蘗鹼對肺癌細胞之生物效應,四株肺癌細胞 H1299、 H1299/p53、A549 以及 CH27 分別種植於 12 孔盤中,分別處理小蘗鹼 1 µM、10 µM 和 100 µM,於顯微鏡下進行觀察。在初期 12 小時內,鏡檢下 觀察並無明顯的差異性,但於給藥 24 小時後,處理組與控制組開始有生 長的速率的不同。48 小時的觀察,可明顯的看出處理組的細胞生長被抑制 (圖 4),但藥物對細胞並無毒殺的效應。到了 72 小時,處理小蘗鹼的細 胞生長抑制作用更趨明顯。然而,細胞仍無受藥物影響而致死的現象產 生,顯示再本實驗處理之濃度範圍內,小蘗鹼對肺癌細胞的影響,並不會 誘使細胞的凋亡或壞死,而是抑制細胞的分裂增殖。. 二、不同劑量小蘗鹼的生長抑制效應: 由圖 4 之結果顯示,小蘗鹼可抑制肺癌細胞增生,因此將四株肺癌細 胞種植於 12 孔盤中,分別於給藥後 12、24、48 和 72 小時後,利用血清 計數器計算出確實的細胞數目,而得一細胞生長的折線圖。由圖 5 得知, 小蘗鹼對四株肺癌細胞均有顯著並相近的生長抑制作用,細胞在處理小蘗 鹼 100 µM 的濃度下,經過 72 小時的培養,發現細胞的增殖現象完全的被 抑制下來,在 10 µM 濃度下也約有 50 至 70%的抑制生長效果;至於在 1 µM 的情況下,只有約 10%的抑制能力。由此可知,小蘗鹼抑制肺癌細胞生長 的作用呈現著劑量與時間依存性的關係,另一方面,由圖 5 的數據可推論, 小蘗鹼為一抑制細胞增生的化合物,不具有細胞毒殺的作用。因即使在 100 µM 的作用濃度下,生長曲線並無下降的趨勢而保持水平。因此,後續的 研究工作,我們以最有效濃度 100 µM 的小蘗鹼來進行實驗。. 36.
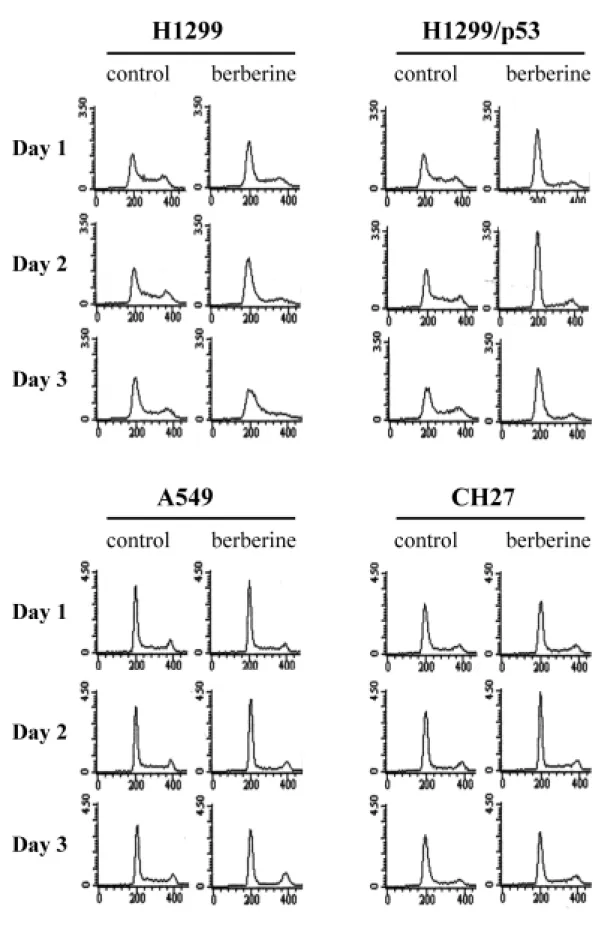
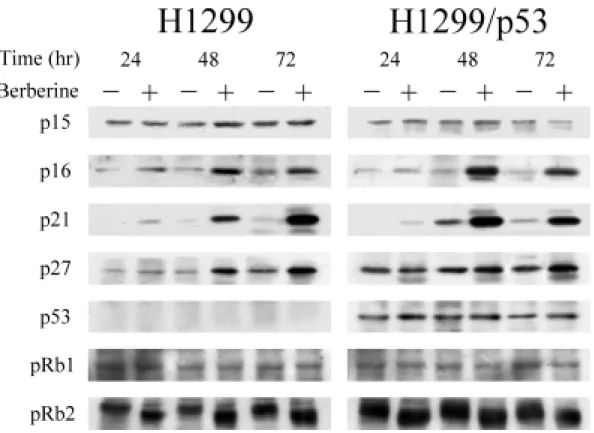
(46) 三、小蘗鹼使細胞週期停滯於 G1 期: 過去的研究已明確的指出,細胞的增生與分裂遵循著一套規律的法 則,稱之為細胞週期。整個週期可分為 G1、S、G2 及 M 期。由上述實驗 結果得知,小蘗鹼會抑制肺癌細胞的分裂增殖,此現象極有可能是透過阻 止細胞週期的進行。因此,利用流式細胞分析儀,分析小蘗鹼是否會影響 對肺癌細胞週期的進行。 由圖 6 結果顯示,小蘗鹼可使 H1299、H1299/p53、A549 以及 CH27 四株肺癌細胞 G1 期的細胞數目隨著處理時間明顯的增加,顯示小蘗鹼的 細胞生長抑制活性,似乎是使細胞無法進入 S 期,而停留在 G1 期。其中 尤以 H1299 與 H1299/p53 兩株細胞最為明顯(表 3)。所以,往後的實驗 就以這兩株細胞來做進一步的探討與比較。 此外,整個分析圖示中並無所謂 sub-G1 peak (亦稱 pre-G1 or apoptotic peak)的出現,進一步證實小蘗鹼並不會造成細胞的凋亡,而只有生長抑 制的作用。. 四、小蘗鹼對細胞週期相關分子的調控: 一般而言,細胞週期的進行與否是靠著許多分子的參與,彼此間訊息 的傳遞環環相扣且錯綜複雜、有時因為一個分子表現量的差異,可能就產 稱截然不同的結果。因此,對於小蘗鹼所導致肺癌細胞週期停滯於 G1 期, 很有可能是小蘗鹼調控了與 G1 期相關分子的表現。所以下一步我們利用 西方墨點法技術,偵測 G1 期相關分子的蛋白表現量,同時也比較對於有 p53 表現的 H1299/p53 細胞株與沒有 p53 表現的 H1299 細胞株,處理小蘗 鹼後分子層次上之異同。 由於 Cdks 以及 cyclins 為細胞週期進行的重要調節因子,首先檢測小 蘗鹼對它們的影響。如圖 7 所示,因小蘗鹼的影響在 G1 期,因此檢測與 G2 期相關的 Cdk1 和 cyclin B 如預測的並無變化;而對於 G1 期相關分子的 37.
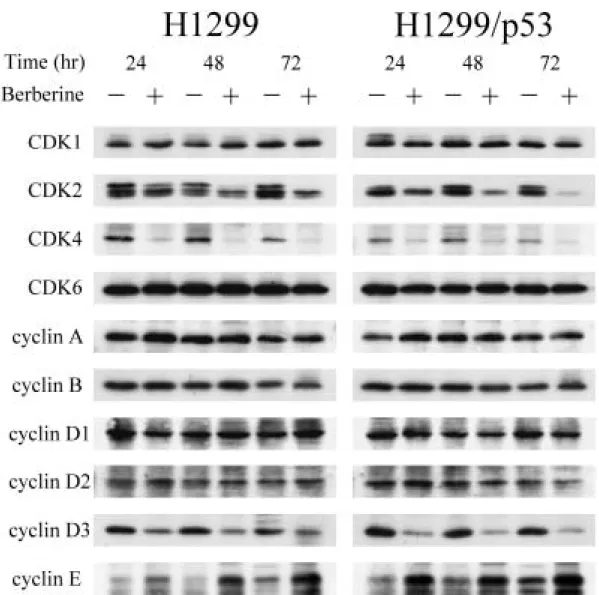
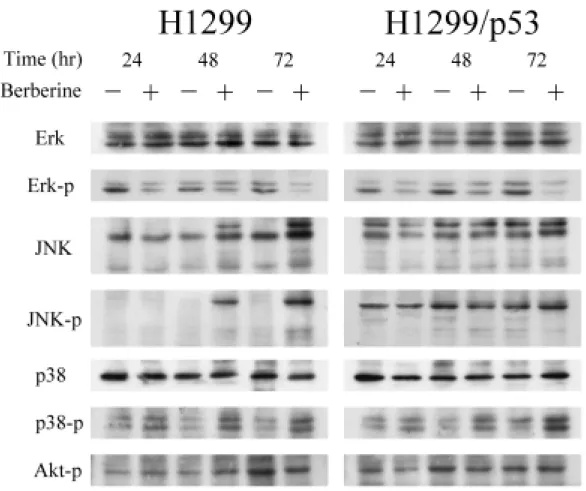
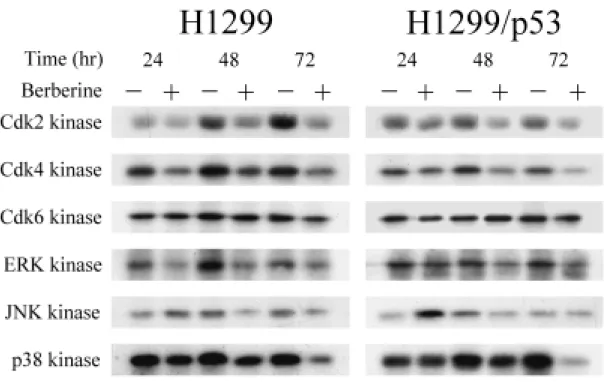
(47) 調控,小蘗鹼在兩株細胞上均造成 G1 早至中期的 Cdk4 和 cyclin D3 以及 G1 晚期相關的 Cdk2 表現量減少,此外,G1 晚期另一相關分子 cyclin E 則 有增加的趨勢,至於 Cdk6、cyclin A、cyclin D1 與 cyclin D2 這些分子則無 變化。 另一方面,探討抑制細胞週期進行的 CdkIs,發現在小蘗鹼的作用下, INK4 家族的 p16 與 CIP/KIP 家族的 p21 及 p27,於兩株細胞的表現量均 有明顯的增加,而 p15 並不受影響(圖 8) 。除此之外,H1299/p53 細胞的 p53 表現量不受調控,同時 H1299 則如預期的偵測不到 p53 表現。另外, 在 Rb protein 家族中的 pRb1 並沒有顯著變化,但是在 pRb2 /p130 蛋白質 表現方面,兩株細胞都可以清楚的看出於給藥組細胞內 pRb2 /p130 蛋白質 磷酸化之程度較低,蛋白質在電泳膠上跑的比較快。而未處理之控制組細 胞中 pRb2 /p130 蛋白質明顯被磷酸化,蛋白質在膠體上移動較慢。. 五、小蘗鹼調控 MAPK 家族分子的活性: 過去的文獻報導指出, MAPK 家族分子參與細胞的生長、分化與死 亡等生命現象之調控,因此,本實驗將探討小蘗鹼是否也藉由調控 MAPK 家族分子去影響細胞的生長,利用西方墨點法技術偵測蛋白質表現量之變 化。由圖 9 的結果得知,於兩株細胞中,ERK、JNK 與 p38 MAPK 三者在 總蛋白質量的表現均無明顯的變化,然而在 ERK 的磷酸化蛋白質表現量 是減少的,但 p38 MAPK 磷酸化蛋白之表現量則顯著增加,至於 JNK 其 磷酸化蛋白質未見有明顯的變化。. 六、小蘗鹼對蛋白質激酶活性的影響: 由以上的實驗結果,許多蛋白質均受到小蘗鹼的調控,其中不乏具有 激酶活性的蛋白質,由於僅靠蛋白質量增減以及磷酸化的有無並不能完全 代表其活性的消長,因此下一步驟便去測定 G1 期分子 Cdk2、Cdk4 和 Cdk6 38.
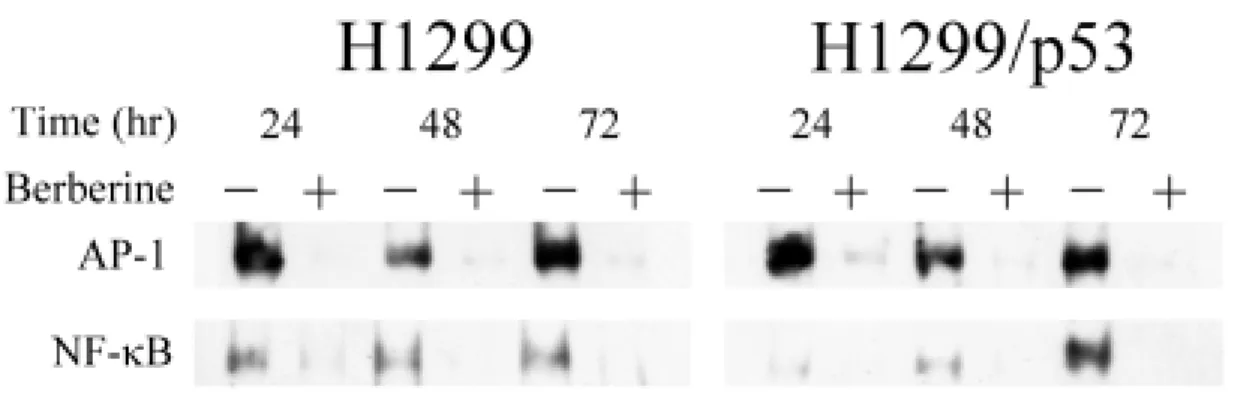
(48) 與 MAPK 家族分子蛋白質激酶活性,是否也隨著蛋白質量的改變而產生變 化。 由圖 10 可明顯的看出,Cdk2、Cdk4 以 ERK 及蛋白質激酶活性與蛋 白質表現量趨勢相同,均有減少的現象,至於 Cdk6 無論於蛋白質激酶活 性或表現量上都沒有太大的變化,然而,p38 MAPK 激酶活性卻是減少的, 與其磷酸化蛋白表現量的趨勢相反,此外,JNK 磷酸化蛋白質雖無明顯的 變化,但其激酶活性於 24 小時處理組為增加的,可是到了 48 和 72 小時 卻反而下降。. 七、小蘗鹼對 AP-1 與 NF-κB 活性的影響: 文獻報導指出,轉錄因子(transcription factors)亦會直接或間接的去 調控細胞週期的進行。其中 AP-1 與 NF-κB 是目前已知與細胞生長、凋亡 以及癌化有極大的關連的轉錄因子,其中 JNK 下游分子中的 c-Jun 為 AP-1 主要組成分子之一,而過去已有研究指出,小蘗鹼可能會抑制 AP-1 的活 性(69)。接著我們利用 Gel Retardation Assay,測定小蘗鹼是否會影響 AP-1 與 NF-κB 和其認知的 DNA 序列結合的能力。一般認為,AP-1 與 NF-κB 如可結合到 DNA 序列上,則可使其調控的下游分子表現量上升,進而影 響一些生物現象。 由圖 11 之結果可知,於兩株肺癌細胞中 AP-1 或 NF-κB 與 DNA 序列 結合的能力均明顯的減少。顯示小蘗鹼可能會透過降低 AP-1 與 NF-κB 與 DNA 結合活性,進而抑制下游分子的表現,此現象可能牽涉小蘗鹼的生長 抑制作用。. 39.
(49) 第五章 討論 小蘗鹼(berberine)為一發現極早的異奎啉類(isoquinoline)植物性 生物鹼,它的蹤跡遍存於眾多科屬的植物當中,而在這些植物當中有不少 為現今常用的中草藥,在過去已有不少學者從事小蘗鹼相關的藥理研究, 顯示其有抗菌和抗發炎等多項的藥理活性,近幾年來,亦有人將小蘗鹼應 用於癌症治療的研究當中,且認為具有發展為抗腫瘤藥的潛力,然其分子 作用機轉仍不甚明瞭,因此,本研究即針對小蘗鹼對非小細胞肺癌細胞的 作用,進行分子層次的探討。 在小蘗鹼處理的四株肺癌細胞–H1299、H1299/p53、A549 和 CH27, 都可見到趨勢類似的生長抑制效果(圖 3) ,比較前人於胃癌細胞 MGC-803 (61). 、食道癌細胞株(YES-1 to YES-6)(63)、血癌細胞 K562(71)以及胃癌細. 胞株 MKN-74、乳癌細胞株 MCF-7 和 MDA-468、結腸癌細胞 HCT-116(62) 等處理小蘗鹼,均顯示出小蘗鹼的抗腫瘤活性是在於其能抑制細胞的生 長,同時可能伴隨著細胞凋亡的現象(61,68,72),然而,於本實驗中肺癌細胞 處理小蘗鹼並無凋亡的現象發生(圖 6) ,有如此的相異點,我們認為應與 各細胞間的分子表現差異性有關,導致各細胞對藥物的敏感度及反應不盡 相同,除此之外,於細胞株 MGC-803 和 YES-1 to YES-6 以及本實驗的四 株肺癌細胞處理小蘗鹼發現可使細胞週期停滯於 G0/G1 期,相反的,於 MKN-74 與 Balb/c 3T3 細胞株所看到的是 G2 期的停止(62,70),顯示小蘗鹼 的抗腫瘤作用機轉可能決定於細胞株的種類。無論如何,由以上之研究結 果,我們可以確定的是小蘗鹼的抗腫瘤活性是透過影響細胞週期正常的運 行。 目前已知,細胞週期可分為 G1 、S、G2 、M 四個時期,當靜止期 (senescence)的細胞接受到 mitogen 的刺激時,細胞便會由 G0 期進入 G1 期,於細胞週期進行期間是受到許多分子的調控,在 G1 的早至中期,cyclin 40.
數據
+6
相關文件
下圖一是測量 1994 年發生於洛杉磯的 Northridge 地震所得 到的圖形。任意給定一個時間 t ,從圖上可看出此時間所對
「世俗化」( secularization)一詞是當下宗教社會學研究中使用
但是 T, A, O, I 出現的次數幾乎不相上下。 要是把每一種組合都試一遍, 直到得出一個 意思 來, 那會是一項沒完沒了的工作。 所以, 只好等新材料來了再說。
1932 年提出李克特量表( Likert Scale ),是一種 心理測量量表,通常用於問卷設計,為目前最受調查 研究者廣泛使用的測量方法.
事實上,就算不是經濟不景,由於現代化下都市生活的發展,經濟和社會情況轉變,對
• 是細胞不正常增生,且這些增生的細胞可
當然,儘管根據以往的經驗,某個問題的解決看似比較容易,但通常事先不會知道困難
這兩棵大榕樹或許生長了幾十年才換來高大的樹杈,深埋在地裡的長
