鞣花酸抑制氧化低密度脂蛋白引發之血管平滑肌細胞增生及凋亡之作用; Ellagic acid inhibit vascular smooth muscle cell proliferation and apoptosis induced by ox-LDL
134
0
0
全文
(2) 1.
(3) 總目錄 頁次 中文摘要 .............................................................................................4 英文摘要 .............................................................................................5 圖目錄 .................................................................................................6 表目錄 ...............................................................................................10 英文縮寫 ........................................................................................... 11 第一章、序論....................................................................................12 第一節、粥狀動脈形成之介紹 ...................................................12 第二節、氧化修飾低密度脂蛋白與自由基的關係......................15 第三節、自由基對平滑肌細胞增生之影響.................................18 第四節、氧化修飾低密度脂蛋白對平滑肌細胞增生之影響.......18 第五節、Ellagic acid 的作用機制...............................................21 第二章、材料與方法.........................................................................24 第三章、實驗結果.............................................................................40 第四章、討論....................................................................................46 第五章、結論....................................................................................52 第六章、序論....................................................................................53 第一節、粥狀動脈形成之介紹 ...................................................53 2.
(4) 第二節、氧化修飾低密度脂蛋白與自由基的關係......................56 第三節、自由基對平滑肌細胞增生之影響.................................59 第四節、ox-LDL 對平滑肌細胞凋亡之影響...............................59 第五節、Ellagic acid 的作用機制...............................................62 第七章、材料與方法.........................................................................65 第八章、實驗結果.............................................................................81 第九章 討論......................................................................................85 第十章、結論....................................................................................92 第十一章、參考文獻.........................................................................93. 3.
(5) 中文摘要 粥狀動脈硬化是一個包含許多功能改變的疾病。氧化低密度脂蛋 白(oxidative low density lipoprotein;ox-LDL)在此疾病中扮演一個 極重要的角色;許多證據證明低密度脂蛋白及氧化低密度脂蛋白皆會 刺激血管平滑肌細胞增生;氧化低密度脂蛋白同時也會引起血管平滑 肌細胞的凋亡。Ellagic acid (EA)是一種存在於蔬果,如:藍莓、 草莓和堅果類中的酚類化合物,可能具有抗氧化、抗癌細胞增生的功 能。因此本論文研究 ox-LDL 刺激平滑肌細胞增生及凋亡與 Ellagic acid 之間的相關性。 本研究使用3-4,5-Dimethylthiazol-2-yl]-2,5-diphenyl-terazolium bromide(MTT)試驗及細胞計數的方式測得細胞之存活率,發現 ox-LDL刺激引起的增生,可經由同步給予Ellagic acid 而有所改善。 流式細胞儀測定細胞週期發現,經ox-LDL刺激平滑肌細胞之細胞週 期S期增加,同步給予Ellagic acid 則可以明顯抑制增生。Ellagic acid 也可以抑制ox-LDL刺激所導致之平滑肌細胞蛋白質,ErK 1/2 、pErK 1/2、和PCNA 的表現增加。在凋亡方面,Ellagic acid 可以抑制細胞 凋亡進行及細胞凋亡時形態的改變,亦可經抑制caspase的基因表現, 進而改善凋亡之情形。這些結果提供食物中的抗氧化劑,可以預防粥 狀動脈硬化疾病的發生。 4.
(6) 英文摘要 In the multifunctional pathogens of atherosclerosis, oxidatively modified low density lipoprotein plays a central role in atherogenesis. Previous studies demonstrated that both native LDL and ox-LDL could stimulate the proliferation of vascular smooth muscle cells. However ox-LDL has also been shown to induce apoptosis in vascular smooth muscle cells (VSMCs). Ellagic acid, an antioxidant, is a phenolic compound presenting in fruits (raspberries, blueberries, strawberries) and walnuts. In this study, we investigated whether VSMCs proliferation and apoptosis induced by ox-LDL would be affected by ellagic acid. Cell viability was tested by MTT test and cell count and study ox-LDL stimulated cell proliferation was inhibited by Ellagic acid. Ellagic acid suppressed the increased the percetage of cell cycle S phase induced by ox-LDL. The activitives of ErK 1/2 , pErK 1/2 and PCNA were also inhibited by ellagic acid. In apoptosis, ellagic acid also inhibited cell apoptosis and cell morphologic change. However ellagic acid suppressed expression of caspase genes could by ox-LDL. These results might significantly advance the understanding of the antioxidants implicated in the formation of atherosclerosis, and could eventually lead to the development of new dietary supplements for the prevention of atherosclerosis. Key word: Ellagic acid, Ox-LDL, Smooth muscle cell, Proliferation, Apoptosis.. 5.
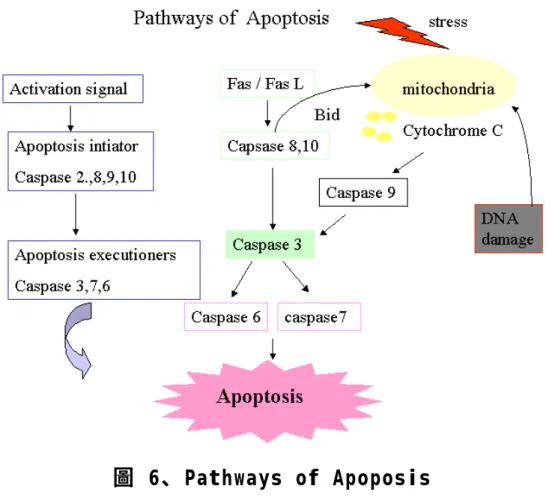
(7) 圖目錄 圖 1、粥狀動脈硬化治病機轉 .......................................................99 圖 2、自由基形成機制圖............................................................. 100 圖 3、MAPK 訊息傳遞途徑............................................................ 101 圖 4、細胞週期及其控制蛋白(cell cycle and control protein) ...................................................................................................... 102 圖 5、Ellagic acid 抑制平滑肌細胞增生可能機制..................103 圖 6、Pathways of Apoposis .....................................................104 圖 7、Ellagic acid 抑制血管平滑肌細胞凋亡可能機制 ...........105 圖 8、以 MTT 評估 Ellagic acid 對血管平滑肌細胞毒性之影響 ................................................................................................106 圖 9、以細胞計數方式評估 Ellagic acid 對血管平滑肌細胞毒性 之影響 .....................................................................................107 圖 10、以 MTT 方式評估低密度之蛋白刺激平滑肌細胞? 生之影響 ............................................................................................... 108 圖 11、以細胞計數方式觀察不同時間點給予 Ellagic acid 對於抑 制低密度脂蛋白刺激血管平滑肌細胞增生之影響 ................ 109 圖 12、以 MTT 方式評估 Ellagic acid 抑制低密度之蛋白刺激平滑 肌細胞? 生之影響................................................................. 110 圖 13、以西方墨點轉漬法觀察低密度脂蛋白對血管平滑肌細胞立即 6.
(8) 性早磷酸化 ErK 1/2 蛋白之影響 .......................................... 111 圖 14、以西方墨點轉漬法觀察 Ellagic acid 抑制低密度脂蛋白刺 激血管平滑肌細胞立即性早期磷酸化 ErK 1/2 蛋白之影響. 112 圖 15、以西方墨點轉漬法觀察 Ellagic acid 抑制低密度脂蛋白刺 激血管平滑肌細胞 ErK 1/2 蛋白之影響 ............................... 113 圖 16、以西方墨點轉漬法觀察 Ellagic acid 抑制低密度脂蛋白刺 激血管平滑肌細胞 PCNA 蛋白之表現..................................... 114 圖 17、以 MTT 方式觀察以不同濃度氧化低密度之蛋白刺激血管平滑 肌 24 小時後細胞之影響 ....................................................... 115 圖 18、以光學顯微鏡觀察同步給予 Ellagic acid 及氧化低密度之 蛋白 100 μg/ml 24 小時後對於血管平滑肌細胞之影響..... 116 圖 19、以 MTT 方式觀察同步給予 Ellagic acid 及氧化低密度之蛋 白 100 μg/ml 24 小時後對於血管平滑肌細胞之影響......... 117 圖 20、以細胞計數方式觀察同步給予 Ellagci acid 及氧化低密度 之蛋白 100 μg/ml 24 小時後對於血管平滑肌細胞之影響. 118 圖 21、以西方墨點轉漬法觀察同步給予 Ellagci acid 及氧化低密 度之蛋白 100 μg/ml 24 小時後對於血管平滑肌細胞之 ErK 蛋 白之影響................................................................................ 119 圖 22、以西方墨點轉漬法觀察同步給予 Ellagic acid 及氧化低密 7.
(9) 度之蛋白 100 μg/ml 24 小時後對於血管平滑肌細胞之早期立 即性磷酸化 ErK 蛋白之影響.................................................. 120 圖 23、以流式細胞儀方法觀察同步給予 Ellagic a cid 及氧化低密 度之蛋白 100 μg/ml 24 小時後對於血管平滑肌細胞週期之影 響........................................................................................... 121 圖 24、以西方墨點轉漬法觀察同步給予 Ellagci acid 及氧化低密 度之蛋白 100 μg/ml 24 小時後對於血管平滑肌細胞之 PCNA 蛋白之影響............................................................................ 122 圖 25、氧化低密度之蛋白刺激平滑肌細胞凋亡之結果.............. 123 圖 26、以細胞計數方式觀察不同時間點給予 Ellagic acid 對於抑 制低密度脂蛋白刺激血管平滑肌細胞增生之影響 ................ 124 圖 27、以 MTT 方式觀察同步給予 Ellagic acid 及氧化低密度之蛋 白 300μg / ml 24 小時後對於血管平滑肌細胞之影響....... 125 圖 28、以細胞計數方式觀察同步給予 Ellagic acid 及氧化低密度 之蛋白 300 μg/ml 24 小時後對於血管平滑肌細胞之影響. 126 圖 29、觀察同步給予 Ellagic acid 及氧化低密度之蛋白 300 μg/ml 24 小時後對於血管平滑肌細胞之影響..................... 127 圖 30、以流式細胞儀方法觀察同步給予 Ellagic acid 及氧化低密 度之蛋白 300 μg/ml 24 小時後對於血管平滑肌細胞凋亡之影 8.
(10) 響........................................................................................... 128 圖 31、以 Giemsa stain 方法觀察同步給予 Ellagic acic 及氧化 低密度之蛋白 300 μg /ml 24 小時後對於血管平滑肌細胞細胞 凋亡型態之影響 .................................................................... 129 圖 32、同步給予 Ellagic acid 及氧化低密度之蛋白 300 μg/ml 24 小時後對計數 Giemsa stain 凋亡之血管平滑肌細胞數....... 130 圖 33、以 MPCR 方法觀察同步給予 Ellagic acid 及氧化低密度之 蛋白 300 μg/ml 24 小時後對於血管平滑肌細胞 Caspase 基因 表現之影響............................................................................ 131. 9.
(11) 表目錄 表格 1........................................................................................... 132 表格 2........................................................................................... 133. 10.
(12) 英文縮寫 ErK 1/2 :Extracellular signal-regulated kinase 1/2 EA:Ellagic acid ICAM-1: Intercellular adhesion molecule-1 LDL:low density lipoprotein MAPK :Mitogen activated protein kinase ox-LDL:oxidized low density lipoprotein PCNA :Proliferating cell nuclear antigen ROS:reactive oxygen species TBARS :Thiobarbituric acid reactive substances VCAM-1:Vascullar cell adhesion molecule-1 VSMCs :Vascular smooth muscle cells. 11.
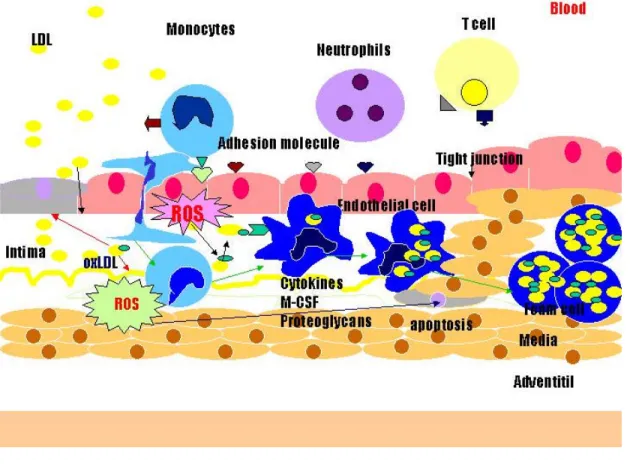
(13) 第一章、序論 前言 任何原因引起的動脈壁增厚、變硬或失去彈性的的病理變化,我們統 稱為動脈硬化(Atherosclerosis)。動脈硬化可依據其硬化方式分為: 動脈粥樣硬化、動脈中層鈣化及小動脈硬化;臨床上以動脈粥樣硬化 為主。粥狀動脈硬化是一種發生在大血管的疾病,是一種脂質與發炎 細胞的聚積,並伴隨著平滑肌細胞(smooth muscle cell, SMC)增生 與細胞外間質液分泌(extracellular matrix secretion),所引起的細胞 內膜纖維變性(intimal fibrosis)。後期血管平滑肌凋亡,所引發粥狀 動脈腫的破裂,栓子及血栓形成,也是導致心血管疾病及中風的主要 原因。根據衛生署公佈民國九十一年十大死因,腦血管與心臟疾病僅 次於惡性腫瘤之後,分居於第二及第三名,不但對個人造成生命財產 上損害,亦花費許多的醫療和社會成本。所以,對於心血管疾病的預 防與治療方法的改進和研發,是一個急待解決及研究的課題。 第一節、粥狀動脈形成之介紹 粥狀動脈硬化形成的機制,主要包括四個階段[1, 2],分別為(1) 內皮細胞層功能不良(endothelial dysfunction)[3, 4]、(2)脂肪條的形 成(fatty-streak fomation)、(3)壞死中心與纖維帽(necrotic core and Fibrous-cap)的形成及(4)不穩定的纖維斑塊(unstable fibrous 12.
(14) plaques)。這四個階段都是影響粥動脈硬化疾病嚴重度的重要的因 數。因此以下將分別介紹各階段: 一 、 內 皮 細 胞 層 功 能 不 良 ( endothelial dysfunction) 早期粥狀動脈硬化形成的原因,主要是因為血管分支或轉彎的地 方,較易為血流速度、壓力或是所產生的擾流影響,使內皮細胞物理 因素改變,發炎細胞[5]的黏著堆積而釋放出趨化因數,導致細胞通透 性增加;加上血液中過多的脂蛋白(例如:低密度脂蛋白;LDL),經 由血液動力學的影響,被動的經由通透性增加的內皮細胞間隙,穿過 血管內皮細胞層,而進入到血管內膜中。這些進入血管壁中的 LDL 會 與結構性蛋白相黏合,且極易受到血管內皮細胞所釋放之活性氧與 12-LO (12/15lipoxygenase) 攻擊,而氧化為氧化態的低密度脂蛋白 (ox-LDL)[6]。 二 、 脂 肪 條 的 形 成 ( fatty-streak formation) 在動脈硬化的第二個階段中,被自由基攻擊的低密度脂蛋白氧化 為氧化態的低密度脂蛋白(ox-LDL),而 ox-LDL 會刺激血管內皮細 胞產生許多的黏附趨化因數,例如 MCP-1(monocyte chemotactic protein)、 ICAM-1(Intercellular adhesion molecule-1),VCAM-1 (vascullar cell adhesion molecule-1)與 E-selectin 等,同時 ox-LDL 亦會吸引血液中的單核球細胞(monocyte)向內皮的表面黏附而進入血 13.
(15) 管內皮細胞下層(subendothelial) ,一方面分泌細胞激素 (cytokines), 如 Interlukin-1(IL-1),Interlukin-4(IL-4),interfiron-? (幹擾素-?; IFN-?),Tumor Necrosis Factor (TNF)進行發炎反應[5, 7];另一方面則 是接著受到血管內皮細胞所釋放的 M-CSF(marophage colony-stimulating factor)分化為巨噬細胞。這些存在於血管壁中的巨 噬細胞表面具有清道夫受體(scavenger receptor) ,對於 ox-LDL 具有 極高的親合力,形成一含巨噬細胞之泡沫細胞(macrophage foam cell) 也稱為”泡沫細胞”。此時平滑肌細胞會受到泡沫細胞的刺激,穿過彈 性纖維層進入內膜增生[8],然後泡沫細胞及平滑細胞,則由內層移位 至內膜增生形成脂肪線(Fatty-streak)[7]。 三 、 壞 死 中 心 及 纖 維 帽 的 形 成 ( necrotic core and Fibrous-cap) 第三階段則由於巨噬細胞不斷的吞噬 ox-LDL,產生大量的泡沫 細胞,極易堆積在血管壁並且死亡[9]。並藉由 Interferon 的作用,使 細胞走向凋亡,再與其他脂質碎片形成一個壞死中心(necrotic core)。血管壁中層膠原蛋白與平滑肌細胞過度增生,遷移至血管內 膜層形成纖維帽(Fibrous-cap),其功能在於隔開血管腔以及受損區 域。 四 、 不 穩 定 的 纖 維 硬 斑 ( unstable fibrous plaques) 動脈硬化最後一個階段中,血管壁會形成動脈硬化斑塊。動脈硬. 14.
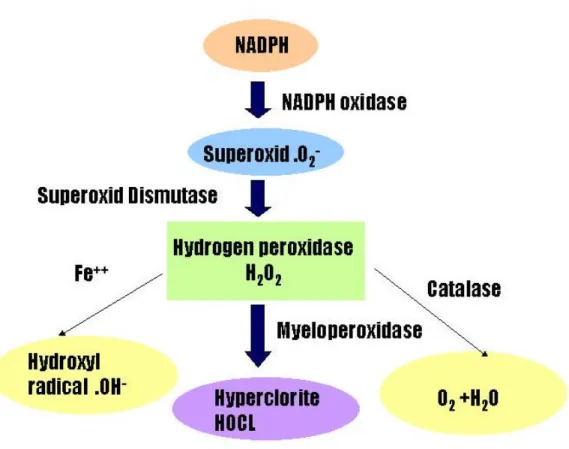
(16) 化斑塊是由平滑肌、泡沫細胞、低密度脂蛋白及其他脂質所組成,也 可以稱為粥狀動脈腫(atheroma)。此時血管的管腔會因為硬化斑塊的 產生而狹窄,持續擠入活化的巨噬細胞會釋放出分解蛋白的酵素,這 些酵素會分解基質,使原本覆蓋於壞死中心上的平滑肌細胞亦受到刺 激進行凋亡之機制[9]。而此機制結果會導致斑塊不穩定,甚至於破 裂,增加粥狀動脈疾病之嚴重度。 第二節、氧化修飾低密度脂蛋白與自由基的關係 低密度脂蛋白(low density lipoprotein, LDL)密度約 1.019-1.063g/ml(Gotto 1988),是由大約 3000 個脂肪酸和一個脫輔 基蛋白分子(Apolipoprotein-B, Apo-B, 50 Dalton)所構成的,平均直 徑長 19-25 nm,分子量大約為 180-250 萬 Dalton 的大圓球形粒子 (Keaney 1994),一般 LDL 被細胞攝取的路徑是經由 LDL 接受體 (LDL receptor)進而分解代謝及應用。 而自由基是指在電子軌域中具有一個或多個不對稱的分子、原子 或離子,則稱之為自由基。當一個分子有不對稱的電子圍繞在外層軌 域上時,會迅速的與周圍的分子碰撞而引起化學變化,進而引起傷害 [10]。在正常情況下,自由基能保護身體免受微生物、細菌等有害物 質侵害,但是當自由基過量時,情況就大不一樣,它會攻擊細胞,促 使蛋白質、核? 酸、脂肪的代謝異常,細胞? 生突變而引起疾病。 體內細胞所產生的活性氧(reactive oxygen species, ROS),是一群 有強氧化力的含氧分子,包括超氧化陰離子(superoxide anion, 15.
(17) O2-),過氧化自由基(hydorxyl radical,OH-)、以及過氧化氫 (hydrogen peroxide)等[11]。自由基具不成對電子其化學活性很活 潑、不穩定,很容易去找一個電子配對,使之穩定,因此產生氧化還 原反應[10]。生物細胞內多種氧化反應及代謝途徑都會產生具有活性 的氧分子,因此生物體常會遭受到活性氧分子之威脅。當生物體老化 或環境惡劣時,粒線體電子傳遞鍊進行不順利時,其電子會在途中被 氧分子作用形成超氧自由基[12]。若超氧自由基與過氧化氫自由基同 時存在,則會產生毒性更強的氫氧自由基。自由基包括超氧自由基 (Superoxide, O2-)、過氧化氫自由基(Hydrogen peroxide, H2O2)、 氫氧自由基(Hydroxyl radical, OH–)、過氧化脂質(Lipid peroxide)。 氧化低密度脂蛋白的形成與由自由基引發的脂質過氧化(lipid peroxidation)有相當的關聯。脂質過氧化是一種自由基的連鎖反應 (chain reaction)。這種連鎖反應是透過羥自由基(hydroxyl radical, OH-),攻擊多元不飽和脂肪酸(polyunsaturated fatty acid,PUFAs) 上的雙鍵(double bonds),羥自由基會吸引多元不飽和脂肪酸碳原子處 的氫原子,先形成不飽和脂肪酸自由基(R.),再氧化成脂過氧基 (ROO.) ,最後生成過氧化脂質(ROOH)及另一份子不飽和脂肪酸自 由基(R.) ,連鎖循環下去,使脂質不斷氧化,生成過氧化脂質。Jessup 16.
(18) et al. (1990)指出 LDL 氧化主要是經由一個自由基反應的過程[13],其 脂質被過氧化作用 (lipid peroxidation) 轉換成脂氫過氧化物,之後再 被分解成不同的產物,包括 malondialdehyde (MDA) , 4-hydroxynonenal 等[14],脂質過氧化的產物會與 LDL 的 apolipoprotein B 作用,致使 apolipoprotein B 的結構受到改變[15]。這 種受到氧化的 LDL(ox-LDL)不再被 LDL 接受體所辨識,而與清道 夫受體有極高的親合力。Rosenfeld et al. (1990)認為 lipid peroxidation 和 lipid peroxidation 的產物在導致粥狀動脈瘤之啟始作用過程中,扮 演著一個重要的角色[16]。另外 ox-LDL 與 LOX-1 結合會減少內皮細 胞內一氧化氮(nitric oxide, NO)的生成,增加 Superoxid 產生[17],而 高膽固醇血症(hypercholesterolaemia)會在內膜產生更多的 ox-LDL, 或在易氧化的環境下,使內皮細胞釋放出更多的超氧陰離子 (superoxide anion, O 2-),結果則造成在沒有明顯傷害時也可能會造成 血管痙攣或收縮。Ox-LDL 會改變動脈壁的特性而造成臨床上之疾 病,會刺激組織釋出凝血因數,並造成凝血 (initiate coagulation)。因 為血管受傷處之週圍,會產生斑塊破裂(plaque rupture),造成栓塞。 在傷害處有許多含有 ox-LDL 之泡沫細胞。因此除了巨噬泡沫細胞之 形成以外,ox-LDL 也是促成粥狀動脈硬化之因素[7]。. 17.
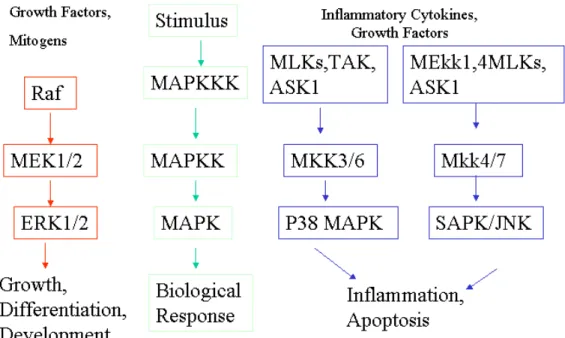
(19) 第三節、自由基對平滑肌細胞增生之影響 增 生 (Proliferation) 和 細 胞 凋 亡 (apoptosis)是血管平滑肌細胞 (vascular smoth muscle cells;VSMCs)造成粥狀動脈硬化、血管再阻塞 及高血壓的兩個重要因素。Li et al. (1997) 指出不同 ROS 對 VSMCs 會有不同的影響,例如超氧陰離子會造成細胞增生;H2O2 會引起 apoptosis[18]。因此 ROS 會參與粥狀動脈硬化之形成,而粥狀動脈硬 化在 O2- 及 H2O2 的影響下,刺激增生和引發 VSMCs 的凋亡。近年來 也有許多文獻指出氧化低密度脂蛋白與細胞增生[19]、細胞毒性[20, 21]及細胞凋亡[22]與低密度脂蛋白過氧化脂質發展過程中的脂質氧 化代謝的產物有關。 第四節、氧化修飾低密度脂蛋白對平滑肌細胞增生之影響 細胞內的 MAPK (Mitogen activated protein kinase pathway)訊息傳 導路徑,在於調控正常細胞功能扮演非常重要的角色。其主要參與細 胞的生長、發育、分化、發炎反應與細胞凋亡等細胞生理反應。當細 胞受到外界如生長因數、細胞激素或壓力(stress)等的刺激下,會誘發 特定訊息傳導路徑中的蛋白質磷酸化,而產生活化作用,並將一系列 訊號,依序的逐漸往細胞核內傳遞,進而產生相對的細胞生理反應。 Yang, et al.等學者指出,在小狗血管平滑肌細胞給予氧化低密度脂蛋 白,會活化 MAPK 中 Erk1/2 訊息傳遞路徑[23]。另外原生(Native) 與氧化(oxidative)的低密度脂蛋白(low density lipoproteins)會經 18.
(20) 由氧化還原反應,間接活化 ERK1/2 MAPK 進而引起平滑肌細胞的增 生[24],楊春滿教授也在 2003 年 Lipid research 中發表的研究報告指 出,氧化低密度脂蛋白會經由 PI3-kinase/Akt 間接活化 MAPK,導致 血管平滑肌細胞的增生[25]。一般而言 MAPK 路徑傳導可被歸納為三 條,第一條是活化 Erk 1/2 為主的傳導路徑,此路徑主要是負責和細 胞生長與分化相關的作用。而第二及第三條則分別為 p38 MAPK 路徑 和 SAPK/JNK 路徑,這兩條路徑則與發炎反應和細胞凋亡有較大之關 連性[26, 27]。 增生除了與MAPK訊息傳遞途徑有關係外,也與細胞週期的進行 有著密切的關係。當細胞藉由生長與分裂間,週而復始的循環,形成 細胞週期(cell cycle),而細胞週期依照其時間順序可分為四期,依 序分別為G1 期、S 期、G2 期及M期,其中S 期為DNA 合成期 (synthesis),而M 期則為細胞有絲分裂期(mitosis)。一般狀態下細胞 多長期處在G0 期或稱為靜止期。但當細胞受到外界增生性刺激因數 的促進下,細胞就會進入到細胞週期之G1期。G1期為DNA 進行合成 前之準備期,此期有一檢查點(check point)或稱為限制點(restriction point)的機制,對於細胞是否繼續進行DNA合成或細胞有絲分裂,做 有效控管的動作。當細胞不適合進行分裂時(例如周遭生長環境不佳 或細胞本身的DNA 受損等),此機制即會終止細胞週期繼續進行。細 19.
(21) 胞在通過G1 和S 期間的限制點機制後,細胞即會進行DNA 合成及 接下來的有絲分裂期,使原來的細胞由一個分裂成兩個細胞而完成一 次完整的細胞週期。細胞週期的進行受大兩股力量之間的調控,一股 是促進細胞週期性的力量,例如來自生長因子、致癌基因 (oncogenes)、CDKs 與Cyclins 等的作用,另一邊則來自例如抑癌基 因或CDK 抑制劑之抑制細胞週期進行的作用。當促進性的力量大於 抑制性的力量,則會促使細胞週期開始運轉,反之則抑制進入細胞週 期。細胞週期限制點的機制主要是以Rb 蛋白質為中心,經由此蛋白 質磷酸化程度來達到調控作用。正常未磷酸化狀態下的Rb 會與稱為 E2F 的轉錄因數結合,當Rb 蛋白質受到活化的cyclin D/CDK4、 cyclinD/CDK6 或cyclin E/CDK2 複合體等的作用而磷酸化後,會促 使與Rb結合的E2F 釋放出來,緊接著E2F 會結合到與細胞週期之S 其相關的基因之特定DNA 序列上,促進這些基因的轉錄作用,進而 使細胞週期由G1期進入S 期和有絲分裂期。這些促進Rb 磷酸化的 cyclin/CDK複合體活性受到兩類蛋白質族群的抑制,一個為Cip/Kip 家族,另一個為Ink 4 家族,而這兩類蛋白質族群被統稱為CDK 抑 制劑(CDKinhibitor; CDKI),並分別對不同種類的cyclin/CDK 複合體 進行抑制的作用[28]。其中Ink 4 家族包含有p14、p15、p16、p18 和 p19 等蛋白質,Cip/Kip 家族則包含p21、p27 和p57 等蛋白質。因 20.
(22) 此如果為了要抑制細胞過度的增生,則可藉由提升CDK 抑制劑的活 性或降低cyclin/CDK 複合體之表現量或活性,即可進進一步抑制Rb 蛋白質的磷酸化,使得E2F 這類促進S 期進行所需的轉錄因數無法 釋放出來而發揮作用,最後達到抑制細胞增生的作用。而氧化低密度 脂蛋白磷脂質結構上的lysophosphatidylcholine ,藉由氧化依附性機 轉,驅使平滑肌細胞進入細胞週期,合成DNA,接著導致平滑肌細胞 釋放出纖維母細胞生長因子,促進平滑肌細胞增生[29, 30]。 第 五 節 、 Ellagic acid 的 作 用 機 制 近幾年來,發現許多存在於自然界的多酚類化合物( polyphenolic compounds)具有多項功能,其中包括抗氧化( antioxidants)[31, 32]、 抗突變(antimutagens) 、抗致癌物(anticarcinogens)及化學保護作用 (chemoprevention )[33, 34],而 Ellagic acid 就是其中一種重要的多 酚類化合物。其化學結構如下: 2,3,7,8-Tetrahydroxy(1)benzopyrano(5,4,3-cde)benzopyran-5,10-dio ne. Ellagic acid 主要存在於自然界中的蔬菜、水果,尤其以草莓、藍 莓及堅果類含量最為豐富。文獻指出 Ellagic acid 抗癌方式是經由活 21.
(23) 化 P53 及其下游 P21 進而達到抑制癌細胞的細胞週期,進而使癌細胞 凋亡,達到抑制癌細胞增生之效果[35]。許多研究證明,Ellagic acid 具 有抗致癌物的活化之功能[36]。Ellagic acid 在化學保護作用 (chemoprevention)方面:根據 2001 Laura A. 研究報告指出在使用 NBBA 誘發 F344 rat 食道癌症形成,可藉由口服乾燥的黑莓,降低食 道腫瘤形成具有化學保護作用[33]。在抗氧化方面,Ellagic acid 可降 低小鼠肝臟、肺臟的 glutathione 和 glutathione reductase ,降低 NADH - and ascorbate-dependent 脂質過氧化(lipid peroxidation)[37],Hassoun 等學者指出,Ellagic acid(6mg/kg per day)可調控抑制氧化劑 TCDD 引起胚胎及胎盤組織的毒素、氧化傷害(Toxicology)[38],另外 Ellagic acid 也可以明顯的抑制 H2O2 及 Bleomycin 所引起的 DNA 傷害[39]。. 22.
(24) 研究目的 本實驗是以體外實驗(in vitro)為主軸的研究,以模擬粥狀動脈 硬化進行ox-LDL刺激平滑肌細胞增生的實驗。在這個研究中,我們 使用了一種自然界蔬果中含有的多酚類化合物 Ellagic acid ,並研究 此多酚類化合物在以ox-LDL造成的平滑肌細胞增生的過程中所扮演 的角色。 1、Ellagic acid 在以往的研究多以預防和治療癌症及抗氧化方面為 主,然而卻少有關於粥狀動脈硬化方面的研究報告。因此我們針對平 滑肌細胞增生與Ellagic acid之間關聯進行研究。 2、探討Ellagic acid 在抑制血管平滑肌細胞增生所扮演的角色是經由 何種機制導致?. 23.
(25) 第二章、材料與方法 第一節、實驗材料 A 儀器 無菌操作臺(造鑫, Taiwan) 細胞培養箱 (Nuaire, USA) 細胞計數器 (Haemocytometer; Boeco, Germany) 光學顯微鏡 (Motic, Japan) 離心機 (Beckman) 酸鹼值測定計 (C831; Consort, UK) 乾浴槽 (Model 110001; 購自Boekel) 微量天平 (GR-200; A&D, Japan) 去離子水製造機 (Minipore, USA) 超高速離心機(himac CS 120GX) Shaker bath (BT-350; 購自YIH DER, Taiwan) 超音波震盪器 Sonicater (MISONIX,Farmingdale,NY ,USA) Power supply ( Hoefer,San Francisco, CA, U.S.A) ELISA reader(ANTHOS-2020,Salzbrug,Austria) Vortex-genie 2 (SCIENTIFIC INDUSTRIES,NY,USA) SDS-PAGE 電泳槽套組 (Amersham, UK) 蛋白質轉漬電泳套組 (Bio-Rad, USA) 水浴槽 Water bath (TKS,Taipei,Taiwan ) 流式細胞儀(FACSCalibur,BD,USA) 24.
(26) 影像分析儀器(AlphaImagerTM 2200,USA) B 材料 細胞培養皿 (騰達行, Taiwan) 細胞培養盤 (騰達行, Taiwan) PVDF 轉漬膜 (Minipore, USA) 蓋玻片 (Kimble, USA) 載玻片 (Marriefeld, Germany) 冷凍管 (騰達行, Taiwan) 微量離心管 (季勗, Taiwan) 離心管 (季勗, Taiwan) Falcon (汎泰, Taiwan) X-film ( Kodak,USA) Paraffin (購自 American national can) 透析膜 (Amersham,UK) PD 10 (Amersham,UK) C 試劑 Acrylamide/Bis (生工, Taiwan) APS (Ammonium persulfate; USB) Bradford reagent (BIO-RAD, Hercules,California,USA) BSA(Bovine serum albumin)(SIGMA, ST.Louis,MO,USA) Bromophenol blue (USB) DMSO (Dimethyl Sulfoxide; Sigma, USA) DTT (1,4-Dithio-D,L-threitol; GERBU, Germany) 25.
(27) DMEM (Dulbeccco’s Modified Eagle’s Medium; GIBCO, USA) ECL kit (Amersham, UK) Ellagic acid (Sigma, USA) Ethanol (景明化工,Taiwan ) FBS (Fetal Bovine serum; GIBCO, USA) Glycine (AppliChem, Germany) Glyerol (Scharlau, Spain) Hydrochloric acid (Merck, USA) Methanol (景明化工,Taiwan) MTT (3-[4,5-Dimethylthiazol-2-yl]-2,5-diphenyl-terazolium bromide; 購自Sigma, USA) Paraformaldehyde (景明化工, Taiwan) Penicillin-Streptomycin (GIBCO, USA) PI (Propidium iodide; Sigma, USA) Potassium dihydrogen phosphate (Merck, USA) Potassium chloride (Scharlau, Spain) Protein assay-Dye reagent concentrate (Bio-Rad, USA) Protein maker (Amersham, UK) RNase A (Sigma, USA) RT kit(Invitrogen, Carlsbad, CA, U.S.A) SDS (Sodium dodecyl sulfate; USB) Sodium bicarbonate (Merck, USA) Sodium chloride (Scharlau, Spain) 26.
(28) Sodium hydroxide (SHOWA, Japan) Sodium pyruvate (GIBCO, USA) TBA (2-Thiobarbituric acid; Sigma, USA) TCA (Trichloroacetic acid; Sigma, USA) TEMED (N,N,N’,N’-Tetramethyl-ethylenediamine; Pharmacia, Sweden) Tris (Tris(hydroxymethly)-aminomethane; Pharmacia,Sweden) Triton X-100 (USB) Trysin-EDTA (GIBCO, USA) Tween 20 (Pharmacia, Sweden) 脫脂奶粉(安佳, New Zealand) 顯影劑(Kodak, USA) 定影劑(Kodak, USA) 一級抗體: (a). anti-PCNA (Santa Cruz, USA) (b). anti-p-Erk1/2 (Santa Cruz, USA) (d). anti-Erk1/2 (Santa Cruz, USA) (e). anti-bata actin (abcam, USA) 二級抗體 anti-mouse IgG (horseradish peroxidase conjugated; Santa Cruz, USA) anti-rabbit IgG (horseradish peroxidase conjugated; Santa Cruz, CA). 27.
(29) 第二節、實驗方法 1、 Ellagic acid 配 製 方 法 秤取Ellagic acid 粉末30.22 mg,加入細胞培養等級之DMSO溶解, 配製成 0.1 µM、1 µM、10 µM、25 µM、50 µM、75 µM、100 µM等 濃度備用。 2、 低 密 度 脂 蛋 白 的 分 離 將抽出的血液樣品放入真空採血管中,於 37℃浴水槽 1.5hr,置 於 4℃冰箱 1hr,離心( 3000 rpm、4℃、15 分鐘)後血清、血球分離, 取上層血清 3 ml 加入溶液配製 0.9 ﹪NaCl 0.7ml 離心(95000 rpm、 10℃、3.5hr、Acc:5、Dec:7) ,去除 VLDL 後直接取 KBr 粉劑 166.8 mg 加入 3ml 血清,離心(、95000 rpm、10℃、3.5hr、Acc:9、Dec: 7)即可得到低密度脂蛋白[24]。 3、 低 密 度 脂 蛋 白 的 氧 化 取 LDL 1 mg/ml 加入 PBS(no EDTA)及 1 mM 硫酸銅(CuSO 4) 最終濃度為 10 µM,37℃水浴 24hr,加入 0.1ml EDTA 10mM(終止 氧化)最終濃度為 1 mM,將約 2∼4 ml 的 LDL 放入膜內放入裝滿 PBS(no EDTA)透析液瓶中,於 4℃冰箱中 stir,透析 24hr 更換 3 次透析液(1hr、2hr、2hr) ,透析 24hr 後自透析膜取出 LDL(體積約 增加 0.5 ml)或使用 PD10 透析,用 0.22 µm 過濾所得即為氧化低密 28.
(30) 度脂蛋白 oxLDL,經由 TBAR 測得其氧化程度[24]。 4、 平 滑 肌 細 胞 的 培 養 血管平滑細胞株(購自財團法人食品工業發展研究所生物資源保存 與研究中心,生資中心編號:60127,細胞株名稱:A10)來自胎鼠 胸主動脈(the thoracic aorta of DB1X embryonic rat),多應用於血管平 滑肌(vascular smooth muscle cells; VSMCs)實驗模型。 培養基及試劑 (A)無血清培養基:Dulbecco’s Modified Eagle’s Medium ( DMEM)Powder : high glucose , L-glutamien , pyridoxine hydrochloride , 10 mg/L sodium pyruvate , 需另外加入 soudium bicarbante 3.7g 。 (B)胎牛血清(Fetal bovine serum ; FBS) 血清購回後需先水浴 56℃加熱 30 分鐘後使用。(一般培養使用 10 % FBS、實驗進行時使用 1 % FBS、同步化時使用 0 % FBS)。 A. HEPES Buffer Solution (1M)使用劑量 1 % B. Strpotomycin/penicilline 使用劑量 1 % C. Trypsine-EDTA 0.25 % Trypsin with 1mM EDTA-4Na 培養方法 將存放血管平滑肌細胞的冷凍管從液態氮中取出後插上浮板,迅速 放入37℃水浴中使其快速解凍,將解凍之細胞懸浮液從冷凍管中放入 29.
(31) 10 公分培養皿,並加入約10 ml 含10 %胎牛血清之培養基,混合均 勻後放入細胞培養箱(37 ℃、95 % O2、5 %CO2) 中進行培養,每 二天換一次培養基。 次培養 當細胞長約 9 分滿時,則進行分盤。首先將舊的培養廢液抽吸乾 淨,加入 7 ml 1x PBS 緩衝液(pH:7.4) 清洗一次,加入 Trypsin- EDTA 1ml incubate 2 分鐘,加入 1 ml 10% FBS- DMEM 2 ml 中和 Trypsin-EDTA 作用,以抽吸方式將細胞沖散,將培養皿液體吸換至離 心管後加入 10 % FBS- DMEM 共 10 ml ,離心 1500 rpm,5 分鐘, 計算細胞數目後進行日常分盤或實驗用分盤。 冷凍細胞:使用 90 % culture medium + 10 % DMSO。 5、 細 胞 數 目 計 算 先將10 公分培養皿內之培養液吸去,再以約7 ml 的1x PBS 緩衝 液(pH=7.4)小心清洗細胞兩次後加入約1 ml 的1x trypsin-EDTA 置入 培養箱內,待2 分鐘後取出。以輕拍培養皿底部的方式將細胞完全脫 離培養皿懸浮起來。加入1 ml 的培養液中和 trypsin-EDTA 活性,並 用1 ml 的pipette 以緩慢來回抽吸方式將細胞打散。將打散後之細胞 換到15 ml 離心管中,並加入10 ml培養液稀釋混勻。如所含細胞密度 過高,則算出的數目誤差較大,需再加入更多的培養液稀釋。取出100 30.
(32) µl 細胞懸浮液和10 µl typan-blue,在96-well內混合均勻,共取2次。 分別從混合液中各取出10 µl 的細胞懸浮液放在細胞計數盤 (haemocytometer)上下兩個凹槽上,利用蓋上蓋玻片時的虹吸作用將 細胞均勻平均分散於細胞計數區域。在顯微鏡下計數細胞數目(細胞 顏色較亮者為存活的細胞)並算出細胞計數盤內9 大格中2 格(左上角 及右上角區域)之活細胞總數(N 值)。N 值除上4,再乘上1.1(稀釋 倍數)x10(細胞懸浮液總量)x 104 即可得10 ml細胞總數目 (cells/10ml)。. 細胞計數盤與細胞計數區域放大簡圖。 6、 細 胞 週 期 同 步 化 (synchronize) 將細胞分到實驗適合之dish或plates後(例如:6cm dish 細胞數目約 3.5 x105 cells/ well),以含10 % 胎牛血清的培養液培養細胞,並於放 入培養箱前稍微搖晃使細胞分佈均勻。待12∼24 小時細胞適應環境 並貼覆上 plates 穩定後吸去培養廢液,再以1XPBS緩衝液(pH:7.4) 2 ml/well 小心清洗1 次,最後加入2 ml/well 的0 %胎牛血清培養 31.
(33) 液,經24小時後即完成細胞同步化步驟。 7、 細 胞 蛋 白 質 抽 取 將培養於 6 cm dish 的細胞取出,加入 5 ml 1x PBS(pH:7.4)沖洗 2 次,加入 500 µl 的 lysis buffer 反應 3 分鐘,使用細胞刮杓將細胞 刮下並將細胞液體收集至微量離心管放入乾浴加溫器內加溫( 95℃、 5 分鐘) ,隨即放置於冰上 30 秒以上,4℃離心(13000 rpm、10 分鐘) , 取上清液,進行蛋白質定量或冰存於-20℃。 ◎ Lysis buffer 配製方法 組成. 最終濃度. 最初濃度. Tris-Hcl(PH:6.8). 6.25M. 0.5M. 體積 / 重量 5ml. SDS. 2%. 10%. 8ml. DTT DDW. 50mM -. 0.3M -. 6.67ml 20.33ml. 總體積. 40ml. 8、 蛋 白 質 定 量 原理 : 確定蛋白質的體積濃度,並使其均一化,以利實驗之進行。 蛋白質檢量線製作 : 依照下列表格進行檢量線製作 組成. NO.. 1. 2. 3. 4. 5. 6. 蛋白質濃度(ug/ml). 0. 5. 10. 15. 20. 25. 0.1mg/ml BSA (µl) DDW(µl). 0 800. 50 750. 100 700. 150 650. 200 600. 250 550. Baradford(µl). 200. 總體積(µl). 1000. NO. 1 為背景值 32.
(34) Bradford 為蛋白質染劑,有毒性,使用時需戴手套。加入染劑混合均 勻後需反應 5 分鐘在測吸光值 。吸光值測定時每個樣本數需 3 重複, 使其數值穩定。以測定出來的標準品吸光質與蛋白質濃度畫出檢量 線,並求出趨勢線方程式即 R2 值。R2 值需 >0.99 以上的準確度。 使用 Excel 軟體會出蛋白質檢量線 算出趨勢線方程式,經帶入測得之吸光值(y),則可求出蛋白質濃度 (µg/ml)。 樣品蛋白質濃度測定 : 取 10 µl 的樣品與 790 µl 的 DDW 混合,在加入 200 µl 的 Bradford 染 劑,均勻混合 5 分鐘後在 O.D 590 測定吸光值。 將樣品吸光值(y)帶入算出趨勢線方程式,則可算出樣品蛋白質濃度 (x) 9、 蛋 白 質 電 泳 ( SDS-PAGE) 組合鑄膠檯,注入 DDW 確定無滲漏之情形,依照下列配方先配 製下層膠。 組成. 下層膠(10%Separation gel) 一片量 兩片量. DDW. 4.75 ml. 9.5 ml. 1.5M Tris(PH:8.8) 0.5M Tris(PH:6.8). 2.5 ml -. 5.0 ml -. 10%SDS. 100 µl. 40%Acrylamide/bis(29:1). 上層膠(5% Stacking gel) 一片量 兩片量 3.04 ml -. 6.08 ml -. 1.25 ml. 2.5ml. 200 µl. 50µl. 100 µl. 2.5 ml. 5.0 ml. 610 µl. 1.22 ml. 10%APS. 50 µl. 100 µl. 50 µl. 100µl. TEMED. 10 µl. 20 µl. 6 µl. 12 µl. 33.
(35) 下層膠注入造膠檯內約八分滿,剩餘的空間先用 DDW 填滿去除上面 的氣泡並壓平膠之上緣,等待凝固約 30 分鐘,造上層膠,TEMED 則 須等下層膠凝固後將 DDW 倒出後加入,下層膠凝固後,注入上層膠, 插入 comb 等待凝固約 30 分鐘將 comb 取出後先用 DDW 清洗 well, 將膠檯放入電泳槽內裝滿 Running buffer,將 loading sample 先加熱 (95℃、5 分鐘)立即放置於冰上冷卻 30 秒以上,將 Marker( 5∼10µl) 和 sample(18∼24 µl)loading 到 well 內,利用 100V 跑電泳約 2 個 半小時,取出下層膠放入 0.1 % Commassie blue 進行蛋白質染色,或 準備進行蛋白質轉漬至 PVDF 膜 Western Blot 步驟[40]。 10、 西 方 墨 點 法 ( Western blot) 準備用物-PVDF membrane 需先用 Methanol 潤濕 30 秒,將下層 膠自水龍頭下取去下,並將膠體放至轉漬夾上其順序如下: 白底網夾 海綿. 正極(+). Filter paper PVDF membrane 膠體(gel) Filter paper 海綿 黑底網夾. 負極(-). 將轉漬夾合上放入 Transfer box 蓋上蓋子放置冰桶中並加滿冰塊,放 入 4℃冰箱內,Transfer 條件設定 100 valtage,轉印時間 1 小時,將 轉印後的 PVDF membrane 放入 5 % fat-free milk 室溫 bloclking 1 小 時,0.1 % PBST 搖盪 5 分鐘,共 3 次,把 membrane 放入含有一級 抗體的封口袋內, 4℃冰箱搖晃至隔天。接著取出 membrane 放入 34.
(36) 0.1 % PBST 搖盪 5 分鐘,共 3 次,將 membrane 放入含有二級抗體, 室溫搖盪 1 小時,取出 membrane 放入 0.1%PBST 搖盪 5 分鐘,共 3 次後進行暗房底片顯影[40]。 ◎ 暗房底片顯影(壓片) 準備用物:1 ml Pipet、1 ml tip、cassete、鑷子、感光底片、剪刀、 透明膠片、ECL、Developer、Fixer 將 memgrane 浸泡於 ECL(比例 為 1:1)混合液中約 30 秒,將 membrane(正面朝上)放置於透明膠 固定好,剪適當大小的底片,進行曝光。(依 membrane 上冷光亮度 決定曝光時間,數秒至 1 小時) ,曝光後用顯影劑及定影劑洗底片, 後將底片風乾保存並進行分析[40]。 ◎Membrane 保存 將壓片後的 membrane以 0.1%PBST 10分鐘 2次,放入含 0.1%PBST 的密封袋 4℃保存。(可保存 2 星期) ◎ Commassie Blue 染色 gel 先以 DDW 清洗 15 分鐘,共 3 次,加入適當的 Commassie Blue 反應 20 分鐘,加入 destain buffer 搖盪至清楚呈現 band。 ◎ Comassie Brilliant Blue R-250 泡製方法 組成. 35. Comassie Brilliant Blue. 最終濃度 -. 最初濃度 -. 體積 / 重量 0.25g. Methanal. -. 100%. 45ml. Acetic acid. -. 100%. 10ml.
(37) DDW. -. -. 總體積 ◎ Dstain buffer 泡製方法 組成 Methanol Acetic acid DDW. 45ml 100ml. 最終濃度. 最初濃度. 體積 /重量. 10% - -. 100%. 100ml. 100% -. 70ml 830ml. 總體積. 1000ml. 11、 Cell count ◎原理 應用Trypen blue exclusion實驗計數樣品中的細胞數,因活細胞會 排斥trypan blue 染劑,僅死細胞會被染成藍色,因此計數亮色的細胞 數可相對得知其細胞存活率。 ◎方法步驟 將細胞分盤至平面 24well 培養,種植細胞數約 5×104/well,以 10%FBS DMEM1ml 飼養 24 小時後,以 1 x PBS(pH:7.4)1ml 沖洗 1 次,加入 0﹪FBS-DMEM 100 ml 同步化 24 小時,以含不同濃度藥物 或低密度之蛋白或氧化低密度之蛋白培養至時間點,加入 200 µl Trypsin incubation 2 分鐘,加入 10% FBS-DMEM 800ul 並將細胞衝散 均勻,取出 100 µl 細胞懸浮液和 10 µl typan-blue 在 96-well 內混合均 勻,共取 2 次。分別從混合液中各取出 10 µl 的細胞懸浮液放在細胞 計數盤(haemocytometer)計數活細胞數。. 36.
(38) 12、 MTT assay ◎ 原理 主要是依賴粒線體中琥珀酸去氫脢的作用將 MTT 3-(4,5-dimethyl thiazol-2-yl)-2,5-diphenyl-tetrazolium bromide) 的 tetrazolium 轉變為 藍色產物 MTT formazan。MTT 轉變僅在活細胞中進行,並堆積在細 胞內,且 formazan 形成量與細胞數目呈正比,加入 10 % SDS-Hcl 溶 解後,可以由波長約 590nm 知吸光測量並定量。由於細胞還原 MTT 的能力代表細胞粒線體的活性,因此可以做為細胞存活率的一個指 標。【MTT 保存方法:避光,冷藏於 2∼8℃。】 ◎方法步驟 將細胞分盤至平面 96 well 培養,種植細胞數約 1×104/well,以 10% FBS-DMEM 100µl 飼養 24 小時後,以 1 x PBS(PH:7.4)100 µl 沖洗 1~2 次,加入 0﹪FBS-DMEM 100µl 同步化 24 小時,以含不同濃度藥 物的 10﹪FBS-DMEM 100 µl 培養至時間點加入濃度 5 mg /ml 的 MTT 佔 DMEM 量的 l/10,培養 4 小時後再加入 100 µl 10﹪SDS-Hcl (終止反應)隔天測吸光值 OD590 nm 、ref 620 nm。 ◎MTT 溶液的配製 ﹙避光 4℃儲存﹚ 組成. 最終濃度. 初濃度. MTT. 5 mg/ml. --. 體積/重量 250 mg. PBS(pH7.4). --. --. 50 ml. 總體積 37. 50 ml.
(39) Solubilization solution 配製﹙室溫儲存﹚ 組成. 最終濃度. SDS. 10%. Hcl. 0.01M 總體積. 初濃度 -. 體積/重量 10 g. 1M. 1 ml 100 ml. 13、 細 胞 週 期 分 析 法 ※細胞前處理 將細胞分盤至 6 cm dish 培養,種植細胞數約 3.5×105/well 10% FBS-DMEM 2 ml 飼養 24 小時,加入 1x PBS 2 ml 沖洗 1-2 次,然後 加入 0% FBS- DMEM 1ml 同步化 24 小時,再以含 1% FBS-DMEM 加 入實驗設計之液體共 5ml 培養至實驗時間點,以 1x PBS 5 ml 沖洗 1-2 次加入 Trypsine 500 µl incubate 2 分鐘後加入 500 µl 10% FBS-DMEM 以 pipet 將細胞沖洗下來,離心(1500 rpm【300g】、5 分鐘)倒掉上 清液,以 1x PBS 1 ml 加入離心管,離心( 1500 rpm【300g】 、5 分鐘) 倒掉上清液,以 70% 冰酒精固定細胞,以 vortex 以 shack 3 的速度 一滴一滴緩慢滴入酒精 2 ml,-20℃冷藏隔天[41]。 ※上機前細胞處理 離心(1500 rpm【300g】 、10 分鐘)倒掉上清液去除酒精,以 1xPBS 1ml 加入離心管,離心(1500 rpm【300g】、10 分鐘),加入 PI 染劑 避光,室溫下 incubater 30 分鐘,換成 FACON 專用管,進行流式細 胞儀分析。 38.
(40) ◎PI stain 染劑的配製 組成. 最終濃度. 初濃度. 體積 / 重量. PI Triton. 0.4 mg/dl 1%. 2 mg/dl 5%. 5 ml 5 ml. RNase A. 0.1 mg/ml. 2 mg/ml. 1.25 ml. 1×PBS. 總體積. -. 13.75 ml 25 ml. 14、 數 據 分 析 實驗結果以 One way ANOVA 計算分析。數據結果以 mean ± SE 表示各項值。 (圖中*表示與 1% FBS-DMEM 相比,P<0.05;#表示 與低密度脂蛋白或氧化低密度脂蛋白組相比,P<0.05。). 39.
(41) 第三章、實驗結果 第 一 節 、 Ellagic acid 對 血 管 平 滑 肌 細 胞 之 影 響 以MTT 及細胞計數方式評估Ellagic acid 對血管平滑肌細胞毒性之 影響 平滑肌細胞在經過24 小時細胞週期同步化後,在含1 %胎牛血清的刺 激下,先同步給予不同濃度的Ellagic acid 處理24 小時。經由加入 MTT 反應4 小時後,再加入solubilization solution,經隔夜後測定吸 光值,或以Trypan blue 染色後,再以細胞計數器計算總細胞中存活 的數目或結果顯示(圖8、9)濃度時100 µM的濃度會導致細胞的毒性; 而濃度50 µM則對細胞沒有毒性。 第 二 節 、 同 步 給 予 LDL 與 Ellagic acid 對血管平滑肌細胞之影響 以 MTT方 式 評 估 低 LDL刺 激 平 滑 肌 細 胞 ? 生 之 影 響 平滑肌細胞在經過24小時細胞週期同步化後,在含0.1%胎牛血清的刺 激下,同步給予不同濃度(20、60、100和200 µg / ml) 的LDL處理24 小時,經由加入MTT 反應4 小時後,再加入solubilization solution, 經隔夜後測定吸光值。由於細胞還原MTT的能力代表細胞粒線體的活 性,因此可以做為細胞存活率的一個指標數目,最後換算出經不同 LDL濃度處理後之細胞存活率。結果顯示(圖10)隨著濃度的增加細胞 存活率呈現劑量依附性(dose dependent)。 40.
(42) 以細胞計數方式觀察不同時間點給予 Ellagic acid 對於抑制 LDL 刺 激血管平滑肌細胞增生之影響 平滑肌細胞在經過24 小時細胞週期同步化後,在含 0.1﹪胎牛血清 及LDL100 µg/ml的刺激下,同步給予濃度 50 µM的Ellagic acid;或先 給予濃度50 µM的Ellagic acid 處理24小時,再給予LDL100 µg/ml刺激 24小時;或先給予LDL100 µg/ml刺激24小時,再給予濃度50 µM的 Ellagic acid 24小時。以Trypan blue 染色後再以細胞計數器計算總細 胞中存活的數目。結果顯示(圖11),同步給予LDL及Ellagic acid 組 別,比較其他組別與單獨給予LDL刺激之組別,有明顯的抑制細胞存 活率之表現。 以 MTT 的 方 式 評 估 LDL100µg/ ml 同步加入 Ellagic acid 對血管平 滑肌細胞數之影響 平滑肌細胞在經過24 小時細胞週期同步化後,在含0.1﹪胎牛血清及 LDL100 µg/ml的刺激下,同步給予濃度50 µM的Ellagic acid 處理經過 24 小時,經由加入MTT 反應4 小時後再加入solubilization solution, 經隔夜後測定吸光值。結果顯示(圖12) 與LDL組別比較同步加入 Ellagic acid 50 µM 組別有的抑制細胞數之表現但統計上無顯著差異。 LDL 對 血 管 平 滑 肌 細 胞 立 即 性 早 期 磷 酸 化 ErK 1/2 蛋白之影響 平滑肌細胞在經過 24 小時細胞週期同步化後,在含 0.1﹪胎牛血清 41.
(43) 及低密度脂蛋白 100 µg/ ml 的刺激下於不同時間點(分別為 5、10、15、 20、30 分鐘)後收取細胞抽取蛋白質,以西方墨點法分析不同時間 點,細胞中磷酸化 Erk1/2 的表現變化。實驗結果顯示(圖 13)細胞中 的磷酸化 Erk 1/2 在 LDL 刺激後 5 分鐘後明顯表現並持續至 30分鐘。 LDL100 µg/ml 同步加入 Ellagic acid 對血管平滑肌細胞立即性早期 磷 酸 化 ErK 1/2 蛋 白 之 影 響 平滑肌細胞在經過24 小時細胞週期同步化後,在含0.1%胎牛血 清及LDL100 µg/ml的刺激下同步給予50 µM的Ellagic acid,處理24 小 時,以西方墨點法分析Ellagic acid,對於細胞中磷酸化Erk 1/2 表現 變化。實驗結果顯示(圖14)LDL組比較,同步加入Ellagic acid會抑制 細胞中磷酸化Erk1/2 的表現。 LDL100µg/ml 同 步 Ellagic acid 對血管平滑肌細胞 ERK 1/2 蛋白之 影響 平滑肌細胞在經過24 小時細胞週期同步化後,在含0.1%胎牛血 清及LDL100 µg/ml的刺激下同步給予50 µM的Ellagic acid,處理24 小 時,以西方墨點法分析 Ellagic acid,對於細胞中Erk 1/2 表現變化。 實驗結果顯示(圖15) 與LDL比較,同步給予Ellagic acid對未磷酸化 Erk1/2 並沒有抑制作用的表現。 LDL100 µg/ml 同步加入 Ellagic acid 對血管平滑肌細胞 PCNA 蛋白 42.
(44) 之影響 平滑肌細胞在經過24 小時細胞週期同步化後,在含0.1%胎牛血 清及LDL100 µg/ml的刺激下同步給予50 µM的Ellagic acid,處理24 小 時,以西方墨點法分析Ellagic acid,對於細胞中PCNA表現變化。實 驗結果顯示(圖16)與LDL組比較,同步加入Ellagic acid 會抑制細胞中 PCNA 的表現。 第三節、同步給予 ox-LDL 及 Ellagic acid 對血管平滑肌細胞增生之 影響 ox-LDL 對 平 滑 肌 細 胞 增 生 之 影 響 平滑肌細胞在經過 24 小時細胞週期同步化後,在含 1 %胎牛血清的 刺激下,同步給予不同濃度(20、60、100、200 和 300 µg/ml)的 ox-LDL, 處理 24小時候,加入 MTT反應 4小時後再加入 solubilization solution, 經隔夜後測定吸光值。結果顯示(圖 17)在濃度 50、100 µg/ml 時細胞 存活率與 1﹪FBS-DMEM 培養的細胞存活率明顯增加至 161﹪而 300 µg/ml 則有顯著抑制存活率之情形約 71﹪。 以光學顯微鏡、MTT 及細胞計數方式評估 ox-LDL100µg/ml 同步加 入 Ellagic acid 對 血 管 平 滑 肌 細 胞 數 及 存 活 率 之 影 響 平滑肌細胞在經過24 小時細胞週期同步化後,在含1﹪胎牛血清及 ox-LDL100 µg/ml的刺激下,同步給予濃度50 µM的Ellagic acid, 處 43.
(45) 理24小時,以Trypan blue 染色後再以細胞計數器計算總細胞中存活 的數目,或經由加入MTT 反應4 小時後再加入solubilization solution, 經隔夜後測定吸光值。結果顯示(圖18、19、20) 與ox-LDL(100 µg/ml) 組別比較同步加入Ellagic acid 50 µM 組別有明顯的抑制細胞數及存 活率之表現。 Ox-LDL 100 µg/ml 同步加入 Ellagic acid 對血管平滑肌細胞蛋白 Erk 1/2 蛋 白 之 影 響 平滑肌細胞在經過24 小時細胞週期同步化後,在含1%胎牛血 清及ox-LDL100 µg/ml的刺激下同步給予50 µM的Ellagic acid,處理24 小時,以西方墨點法分析Ellagic aicd對於細胞中Erk 1/2 表現變化。 實驗結果顯示(圖21) ox-LDL (100 µg/ml)組比較,同步加入Ellagic acid 會抑制細胞中Erk1/2 的表現。 Ox-LDL100 µg/ml 同步加入 Ellagic acid 對血管平滑肌細胞立即性早 期 磷 酸 化 ErK 1/2 蛋 白 之 影 響 平滑肌細胞在經過24 小時細胞週期同步化後,在含1%胎牛血清及 ox-LDL100 µg/ml的刺激下同步給予50µM的Ellagic acid,處理24 小 時,以西方墨點法分析Ellagic acid對於細胞中磷酸化Erk 1/2 表現變 化。實驗結果顯示(圖22) 與ox-LDL(100 µg/ml)組比較,同步加入 Ellagic acid會抑制細胞中磷酸化Erk1約0.52倍、Erk2約0.49倍的表現。 44.
(46) Ox-LDL100 µg/ml 同步加入 Ellagic acid 對血管平滑肌細胞週期之影 響 將血管平滑肌細胞經過 24 小時無血清培養進行細胞同步化後,置 換為含1 %胎牛血清之培養液,並分別加入ox-LDL 100µg /ml及 Ellagic acid 50 µM或加入DMSO。於第24 小時收集細胞,並經過細胞 酒精固定和PI 染劑染色後,以流式細胞儀進行分析不同濃度藥物處 理,對血管平滑肌細胞之細胞週期S 期比例影響變化。由結果顯示(圖 23、表一)當給予ox-LDL(100 µg /ml)組別細胞週期S 期與1% FBS-DMEM比較有明顯由6.76%增加到15.42%,而給予ox-LDL(100 µg /ml)並同步加入Ellagic acid 組別則細胞週期S期則降至9.4%且具 有統計上的意義P<0.005。 100 µg/ml ox-LDL 同 步 加 入 Ellagic acid 對血管平滑肌細胞 PCNA 蛋白之影響 平滑肌細胞在經過24 小時細胞週期同步化後,在含1%胎牛血 清及ox-LDL100 µg/ml的刺激下同步給予50 µM的Ellagic acid,處理24 小時,以西方墨點法分析Ellagic acid對於細胞中PCNA 表現變化。實 驗結果顯示(圖24) 與ox-LDL(100 µg/ml)組,同步加入Ellagic acid 會抑制細胞中PCNA 的表現。. 45.
(47) 第四章、討論 本研究主要以體外(in vitro)實驗為主軸,模擬粥狀動脈血管疾 病中血管平滑肌細胞受到 ox-LDL 刺激後所引平滑肌細胞增生與天然 蔬果中具有抗氧化功能之多酚類化合物 Ellagic acid 之研究探討。 Ellagic acid 對 ox-LDL 刺 激 平 滑 肌 細 胞 增 生 的 影 響 有許多證據指出在粥狀動脈血管疾病中 ox-LDL 所引起內膜層平 滑細胞遷移及增生對於疾病嚴重度的發展是非常重要的因素[42, 43]。而文獻指出 ox-LDL 的生物活性可以引起生長因數調控基因、黏 著分子表現和改變單核球及巨噬細胞的能力與平滑肌細胞的增生[7, 8, 30]。 近年研究報告對於粥狀動脈血管平滑肌細胞增生的探討我們可 以發現,有許多的因素可以引起血管平滑肌的增生。如 ox-LDL 和 acetyl-LDL 引起巨噬細胞吞噬,形成泡沫細胞[44];另有文獻指出 LDL 與 ox-LDL 可以藉由活化訊息傳遞路徑 MAPK patheway,或藉由 PI3-kinase/Akt 與 ERK1/2 之間的 cross link引起細胞的增生及遷移[24, 30, 44, 45];另外 ox-LDL 也可以引起細胞內生長因數,如 FGF-2 之 釋放,達到刺激細胞增生。Marjorie 等學者提出,ox-LDL 刺激平滑 肌細胞增生,主要是引起細胞週期蛋白質的表現。細胞週期是由許多 不同的蛋白質所共同調控,包括:cyclin D1、cyclin-dependent kinase 46.
(48) (cdks) or PCNA 等[46]。Proliferating cell nuclear antigen(PCNA) 是 DNA polymerased 輔助蛋白,主要分佈於核質,其主要扮演的角色 是與 cyclin 或 cdks 及 P21 相互作用以調控細胞週期的進行並與 DNA 合成有關[46];而 ox-LDL 氧化產物包括 4-hydroxynonenal、 lysophosphatidylcholine 也會刺激平滑肌細胞的增生[19, 47]。 根據文獻指出自由基亦是導致平滑肌細胞增生的主要原因[24]; 而本實驗所使用的 Ellagic acid,過去主要是針對癌症與抗氧方面的領 域研究,對於 Ellagic acid 在粥狀動脈硬化血管疾病的作用機制並不 清楚。粥狀動脈是一個慢性發炎伴隨著自由基傷害、脂質堆積與平滑 肌不正常細胞增生的血管性疾病,不正常的細胞增生就如同癌細胞一 樣。而 Ellagic acid 在抗癌部分可藉由活化 21 進而抑制癌細胞週期導 致細胞凋亡[35],或經由抑制乙醯轉移酵素活性及 Cytochrome P450 的作用預防致癌物產生及腫瘤生成[33]。而在 TCDD 引起的胎兒毒素 (fetotoxicity)及氧化壓力(oxidative stress)之下,使用 100 µg /kg 濃 度的 Ellagic acid 則可有效的預防胎兒死亡(fatal death)和 speroxide anion 的產生[38]。 在平滑肌細胞增生方面之實驗顯示,氧化低密度脂蛋白(ox-LDL) 與 MAPK 訊息傳遞路徑具相關性[23],並加速細胞週期的進行。本 實驗在給予 Ellagic acid 後,則可以抑制細胞週期、細胞增生、Erk 47.
(49) 磷酸化及 PCNA 的表現,表示 Ellagic acid 可以阻斷 MAPK 的訊息傳 遞及細胞週期的進行。而本研究將 Ellagic acid 應用在粥狀動脈血管 平滑肌細胞的增生的模型,並利用低血清的環境進行實驗。主要是血 清本身含有生長因數可以促進細胞增生,若給予正常飼養濃度的血清 10%可能無法看到顯著的細胞增生情形,而血清可能中和 EA 之作用 而無法清楚釐清 EA 功能為何。在實驗中發現在 Ellagic acid 在濃度 1 ∼40 µM的時候,經 MTT 結果顯示細胞的存活率高於 1﹪FBS-DMEM 組別而在濃度 50 µmol /l 的濃度下則無明顯的差異。因此我們選擇 50 µM 為實驗的濃度,結果顯示血管平滑肌細胞在給與 100 µg /ml 氧化 低密度脂蛋白(ox-LDL)處理 24 小時,會促進細胞增生,且持續磷 酸化 MAPK 訊息傳遞路徑下游之 ErK 蛋白質、細胞週期的 S 期及 PCNA 蛋白質。當同步給予濃度 50 µM 的 Ellagic adid 的確可以明顯 的抑制上述的表現,這意味著 Ellagic acid 的確可以藉由抑制 MAPK 訊息傳遞路徑及細胞週期進而抑制氧化低密度脂蛋白引起的平滑肌 細胞增生。Ellagic acid 抑制氧化低密度脂(ox-LDL)所引起的 MAPK 訊息傳遞路徑也許是經由抑制活性氧( ROS)的產生及活化 P21 而達 到抑制平滑肌細胞的增生;另一方面文獻也指出 cytochrome P450 會 促進低密度脂蛋白的氧化[48], Ellagic acid 則可抑制 cytochrome P450 的產生,進而達到抑制脂質的氧化。 48.
(50) Ellagic acid 抗 氧 化 對 平 滑 肌 細 胞 增 生 與 之 討 論 在本論文研究探討實驗中可以發現,ox-LDL 在不同濃度處理 24 小時候會引起血管平滑肌細胞表現出不同的結果,這也說明瞭 ox-LDL 在粥狀動脈血管疾病中所扮演的複雜角色。2000 年亦有文獻 指出,內皮細胞的 ox-LDLreceptor-1(LOX-1)與 ox-LDL 結合後會 產生 ROS,並且隨著 ox-LDL 濃度增加 ROS 的量亦會隨之增加[49], 血管平滑肌細胞亦被發現有 LOX-1 而 ox-LDL 脂質氧化產物 lysophosphatidylcholine (LPC) 會增加血管平滑肌細胞 LOX-1mRNA 的表現[50],推測這個現象的原因可能是因為 ox-LDL 在不同濃度所 產生自由基的總量不同引起不同的細胞外訊息傳遞路徑因而導致增 生的結果;另一方面也可能是 ox-LDL 脂質氧化代謝產物不同或代謝 產物總量的差異引起細胞毒性而導致不同的結果。 抗氧化劑具有保護身體,防止自由基形成、去除體內所產生的自 由基 (freeradicals);這些自由基是由氧氣和體內的各種物質反應而 產生,會造成一連串有害的連鎖反應,而對體內組織造成各種傷害。 抗氧化劑,例如:脂溶性維他命E、水溶性維他命C,雖然抗氧化機 制不同但是對於降低氧化傷害[51-53],及抑制血液中膽固醇含量都有 顯著的成效[54],並且可以降低並預防心血管疾病及粥狀動脈硬化疾 病進展[55]。而本實驗運用不同時間點的方式,探討Ellagic acid抗氧 49.
(51) 化的機制起始為何,發現同步給予24小時處理的Ellagic acid和LDL, 可以明顯的抑制平滑肌細胞的增生;然而先單獨給予EA或LDL雖然 有抑制細胞增生情形,但是卻不如同步給予明顯。LDL本身並不會產 生ROS但是當加入細胞內時則可引細胞內ROS的產生。因此推論 Ellagic acid在抑制LDL引起的細胞增生,主要是經由捕捉細胞內ROS 進而抑制細胞增生。另外,本實驗所使用的Ellagic acid 即是種天然 抗氧化劑在50 µM的濃度下對於LDL或ox-LDL所引起的增生實驗 中,發揮了抑制細胞增生的效果,而這個濃度與2000年Chai學者提 出,抗氧化劑vitamin E在50 µM具有時抑制lyso PC 所引起的平滑肌 細胞增生[56]。Ellagic acid是一種蔬果中的多酚類化合物其,本身具 抗氧化功能就有保護正常細胞免於不正常環境刺激下受到影響,雖然 文獻[22]有指出LDL或ox-LDL會增加ROS的生成進而影響平滑肌細 胞的增生,但本實驗卻欠缺ox-LDL導致自由基增加直接證據,這也 是論文不足的地方,因此未來實驗的方向可朝向ox-LDL與自由基的 相關實驗及Ellagic acid在抗氧化中所扮演的角色。 粥狀動脈血管疾病是高死亡率的現代文明病的一種,許多研究學 者致力於研究其治病機轉,尋找可以預防或降低此疾病的發生率,而 本實驗使用了存在於蔬果內的具抗氧化功能之多酚類化合物進行實 驗。結果顯示 Ellagic acid 在粥狀動脈中平滑肌細胞增生具有抑制作 50.
(52) 用,這也是本研究的主旨,對於粥狀動脈疾病的進展可以藉由日常生 活中的蔬果的補充,而達到預防或降低的效果,更可以減少因服用化 學合成藥物副作用所造成的更進一步的傷害。. 51.
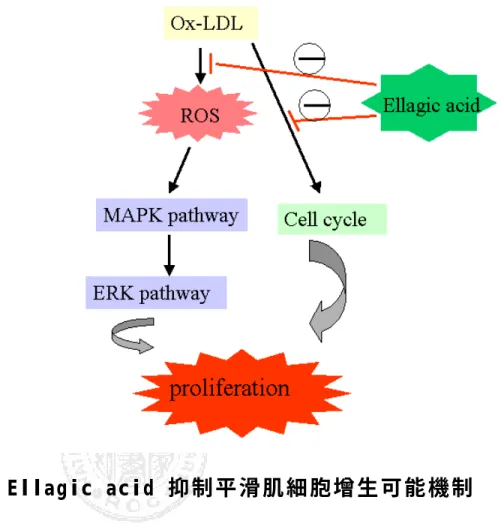
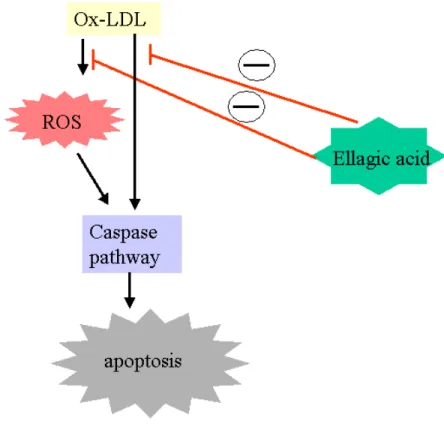
(53) 第五章、結論 存在於自然界的多酚類化合物 Ellagic acid,可以藉由其抗氧化之 功能,抑制平滑肌細胞增生。這個結果可能是經由捕捉 ROS,避免 細胞外的訊息傳遞路徑的活化,或直接抑制細胞週期中蛋白質的表 現,而達到抑制平滑肌細胞不正常增生(圖 5)。Ellagic acid 存在於 自然界蔬果中,或許可以藉由補充蔬果而直接預防或治療粥狀動脈硬 化血管疾病。. 52.
(54) 第六章、序論 前言 任何原因引起的動脈壁增厚、變硬或失去彈性的的病理變化,我們統 稱為動脈硬化(Atherosclerosis)。動脈硬化可依據其硬化方式分為: 動脈粥樣硬化、動脈中層鈣化及小動脈硬化;臨床上以動脈粥樣硬化 為主。粥狀動脈硬化是一種發生在大血管的疾病,是一種脂質與發炎 細胞的聚積,並伴隨著平滑肌細胞(smooth muscle cell, SMC)增生 與細胞外間質液分泌(extracellular matrix secretion),所引起的細胞 內膜纖維變性(intimal fibrosis)。後期血管平滑肌凋亡,所引發粥狀 動脈腫的破裂,栓子及血栓形成,也是導致心血管疾病及中風的主要 原因。根據衛生署公佈民國九十一年十大死因,腦血管與心臟疾病僅 次於惡性腫瘤之後,分居於第二及第三名,不但對個人造成生命財產 上損害,亦花費許多的醫療和社會成本。所以,對於心血管疾病的預 防與治療方法的改進和研發,是一個急待解決及研究的課題。 第一節、粥狀動脈形成之介紹 粥狀動脈硬化形成的機制,主要包括四個階段[1, 2],分別為(1) 內皮細胞層功能不良(endothelial dysfunction)[3, 4]、(2)脂肪條的形 成(fatty-streak fomation)、(3)壞死中心與纖維帽(necrotic core and Fibrous-cap)的形成及(4)不穩定的纖維斑塊(unstable fibrous 53.
(55) plaques)。這四個階段都是影響粥動脈硬化疾病嚴重度的重要的因 數。因此以下將分別介紹各階段: 一 、 內 皮 細 胞 層 功 能 不 良 ( endothelial dysfunction) 早期粥狀動脈硬化形成的原因,主要是因為血管分支或轉彎的地 方,較易為血流速度、壓力或是所產生的擾流影響,使內皮細胞物理 因素改變,發炎細胞[5]的黏著堆積而釋放出趨化因數,導致細胞通透 性增加;加上血液中過多的脂蛋白(例如:低密度脂蛋白;LDL),經 由血液動力學的影響,被動的經由通透性增加的內皮細胞間隙,穿過 血管內皮細胞層,而進入到血管內膜中。這些進入血管壁中的 LDL 會 與結構性蛋白相黏合,且極易受到血管內皮細胞所釋放之活性氧與 12-LO (12/15lipoxygenase) 攻擊,而氧化為氧化態的低密度脂蛋白 (ox-LDL)[6]。 二 、 脂 肪 條 的 形 成 ( fatty-streak formation) 在動脈硬化的第二個階段中,被自由基攻擊的低密度脂蛋白氧化 為氧化態的低密度脂蛋白(ox-LDL),而 ox-LDL 會刺激血管內皮細 胞產生許多的黏附趨化因數,例如 MCP-1(monocyte chemotactic protein)、 ICAM-1(Intercellular adhesion molecule-1),VCAM-1 (vascullar cell adhesion molecule-1)與 E-selectin 等,同時 ox-LDL 亦會吸引血液中的單核球細胞(monocyte)向內皮的表面黏附而進入血 54.
(56) 管內皮細胞下層(subendothelial) ,一方面分泌細胞激素 (cytokines), 如 Interlukin-1(IL-1),Interlukin-4(IL-4),interfiron-? (幹擾素-?; IFN-?),Tumor Necrosis Factor (TNF)進行發炎反應[5, 7];另一方面則 是接著受到血管內皮細胞所釋放的 M-CSF(marophage colony-stimulating factor)分化為巨噬細胞。這些存在於血管壁中的巨 噬細胞表面具有清道夫受體(scavenger receptor) ,對於 ox-LDL 具有 極高的親合力,形成一含巨噬細胞之泡沫細胞(macrophage foam cell) 也稱為”泡沫細胞”。此時平滑肌細胞會受到泡沫細胞的刺激,穿過彈 性纖維層進入內膜增生[8],然後泡沫細胞及平滑細胞,則由內層移位 至內膜增生形成脂肪線(Fatty-streak)[7]。 三 、 壞 死 中 心 及 纖 維 帽 的 形 成 ( necrotic core and Fibrous-cap) 第三階段則由於巨噬細胞不斷的吞噬 ox-LDL,產生大量的泡沫 細胞,極易堆積在血管壁並且死亡[9]。並藉由 Interferon 的作用,使 細胞走向凋亡,再與其他脂質碎片形成一個壞死中心(necrotic core)。血管壁中層膠原蛋白與平滑肌細胞過度增生,遷移至血管內 膜層形成纖維帽(Fibrous-cap),其功能在於隔開血管腔以及受損區 域。 四 、 不 穩 定 的 纖 維 硬 斑 ( unstable fibrous plaques) 動脈硬化最後一個階段中,血管壁會形成動脈硬化斑塊。動脈硬. 55.
(57) 化斑塊是由平滑肌、泡沫細胞、低密度脂蛋白及其他脂質所組成,也 可以稱為粥狀動脈腫(atheroma)。此時血管的管腔會因為硬化斑塊的 產生而狹窄,持續擠入活化的巨噬細胞會釋放出分解蛋白的酵素,這 些酵素會分解基質,使原本覆蓋於壞死中心上的平滑肌細胞亦受到刺 激進行凋亡之機制[9]。而此機制結果會導致斑塊不穩定,甚至於破 裂,增加粥狀動脈疾病之嚴重度。 第二節、氧化修飾低密度脂蛋白與自由基的關係 低密度脂蛋白(low density lipoprotein, LDL)密度約 1.019-1.063g/ml(Gotto 1988),是由大約 3000 個脂肪酸和一個脫輔 基蛋白分子(Apolipoprotein-B, Apo-B, 50 Dalton)所構成的,平均直 徑長 19-25 nm,分子量大約為 180-250 萬 Dalton 的大圓球形粒子 (Keaney 1994),一般 LDL 被細胞攝取的路徑是經由 LDL 接受體 (LDL receptor)進而分解代謝及應用。 而自由基是指在電子軌域中具有一個或多個不對稱的分子、原子 或離子,則稱之為自由基。當一個分子有不對稱的電子圍繞在外層軌 域上時,會迅速的與周圍的分子碰撞而引起化學變化,進而引起傷害 [10]。在正常情況下,自由基能保護身體免受微生物、細菌等有害物 質侵害,但是當自由基過量時,情況就大不一樣,它會攻擊細胞,促 使蛋白質、核? 酸、脂肪的代謝異常,細胞? 生突變而引起疾病。 體內細胞所產生的活性氧(reactive oxygen species, ROS),是一群有強 氧化力的含氧分子,包括超氧化陰離子(superoxide anion,O2-),過氧 56.
(58) 化自由基(hydorxyl radical,OH-)、以及過氧化氫(hydrogen peroxide) 等[11]。自由基具不成對電子其化學活性很活潑、不穩定,很容易去 找一個電子配對,使之穩定,因此產生氧化還原反應[10]。生物細胞 內多種氧化反應及代謝途徑都會產生具有活性的氧分子,因此生物體 常會遭受到活性氧分子之威脅。當生物體老化或環境惡劣時,粒線體 電子傳遞鍊進行不順利時,其電子會在途中被氧分子作用形成超氧自 由基[12]。若超氧自由基與過氧化氫自由基同時存在,則會產生毒性 更強的氫氧自由基。自由基包括超氧自由基(Superoxide, O2-)、過氧化 氫自由基(Hydrogen peroxide, H2O2)、氫氧自由基(Hydroxyl radical, –. OH )、過氧化脂質(Lipid peroxide)。 氧化低密度脂蛋白的形成與由自由基引發的脂質過氧化(lipid peroxidation)有相當的關聯。脂質過氧化是一種自由基的連鎖反應 (chain reaction)。這種連鎖反應是透過羥自由基(hydroxyl radical, OH-),攻擊多元不飽和脂肪酸(polyunsaturated fatty acid,PUFAs) 上的雙鍵(double bonds),羥自由基會吸引多元不飽和脂肪酸碳原子處 的氫原子,先形成不飽和脂肪酸自由基(R.),再氧化成脂過氧基 (ROO.) ,最後生成過氧化脂質(ROOH)及另一份子不飽和脂肪酸自 由基(R.) ,連鎖循環下去,使脂質不斷氧化,生成過氧化脂質。Jessup et al. (1990)指出 LDL 氧化主要是經由一個自由基反應的過程[13],其 57.
(59) 脂質被過氧化作用 (lipid peroxidation) 轉換成脂氫過氧化物,之後再 被分解成不同的產物,包括 malondialdehyde (MDA) , 4-hydroxynonenal 等[14],脂質過氧化的產物會與 LDL 的 apolipoprotein B 作用,致使 apolipoprotein B 的結構受到改變[15]。這 種受到氧化的 LDL(ox-LDL)不再被 LDL 接受體所辨識,而與清道 夫受體有極高的親合力。Rosenfeld et al. (1990)認為 lipid peroxidation 和 lipid peroxidation 的產物在導致粥狀動脈瘤之啟始作用過程中,扮 演著一個重要的角色[16]。另外 ox-LDL 另外 ox-LDL 與 LOX-1 結合 會減少內皮細胞內一氧化氮(nitric oxide, NO)的生成[17],增加 Superoxid 產生,而高膽固醇血症(hypercholesterolaemia)會在內膜產生 更多的 ox-LDL,或在易氧化的環境下,使內皮細胞釋放出更多的超 氧陰離子(superoxide anion, O 2-),結果則造成在沒有明顯傷害時也可 能會造成血管痙攣或收縮。Ox-LDL 會改變動脈壁的特性而造成臨床 上之疾病,會刺激組織釋出凝血因數,並造成凝血 (initiate coagulation)。因為血管受傷處之週圍,會產生斑塊破裂(plaque rupture),造成栓塞。在傷害處有許多含有 ox-LDL 之泡沫細胞。因此 除了巨噬泡沫細胞之形成以外,ox-LDL 也是促成粥狀動脈硬化之因 素[7]。. 58.
(60) 第三節、自由基對平滑肌細胞增生之影響 增 生 (Proliferation) 和 細 胞 凋 亡 (apoptosis)是血管平滑肌細胞 (vascular smoth muscle cells;VSMCs)造成粥狀動脈硬化、血管再阻塞 及高血壓的兩個重要因素。Li et al. (1997) 指出不同 ROS 對 VSMCs 會有不同的影響,例如超氧陰離子會造成細胞增生;H2O2 會引起 apoptosis[18]。因此 ROS 會參與粥狀動脈硬化之形成,而粥狀動脈硬 化在 O2- 及 H2O2 的影響下,刺激增生和引發 VSMCs 的凋亡。近年來 也有許多文獻指出氧化低密度脂蛋白與細胞增生[19]、細胞毒性[20, 21]及細胞凋亡[22]與低密度脂蛋白過氧化脂質發展過程中的脂質氧 化代謝的產物有關。 第 四 節 、 ox-LDL 對 平 滑 肌 細 胞 凋 亡 之 影 響 凋亡(Apoptosis)是一個維持體內環境平衡,及個體生長發 育的一個重要因素[57]。當細胞要進行 Apoptosis 時,會有以下四個 重要外在特徵:1.細胞皺縮(cell membrane shrinkage);2.染色質濃 縮(chromosome condensation)聚集在核膜;3.( DNA fragementaion); 4.凋亡小體(apoptotic bodies)的形成。 細胞的這些改變主要是透過一連串的訊息傳遞和活化 caspase 調控 pro- and anti-apoptosis protein 。例如 Bcl-2 家族成員經由一連 串的正向、負向調控後決定凋亡程式啟動與否。Apoptosis 可經由外 在途徑及內再途徑所引發,外在途徑如死亡接受器(death receptor), 59.
(61) 如;TNFR、Fas、Decoy receptor 和 death receptor[58, 59]。這些 receptor 不管細胞外、 ligand 結合區域和細胞內死亡區域(cytoplasmic death domain) 都富有 cystein,當 ligand 結合上這些接受器(receptor)及死亡 區 域 (death domain)後 會 和 adaptor protein 相 互 作 用 進 而 活 化 caspase。內在途徑凋亡程式啟動則可藉由 Bcl-2 protein 家族成員對 於 apoptosis 的調控或經由 DNA 損傷或基因調控粒腺體釋放出 cytochrome C 進而活化 caspase [60, 61]。 Caspase 家族基因與細胞凋亡程式的進行具有相當大的關係 [62]。不管是內在或外在途徑,最後都會活化 Caspase 進而產生細胞 凋亡。Caspase (aspartate-specific cystein protease) 在 apoptosis 一連串 的過程中扮演起始及終結者之角色 [63]。所謂 caspase 起始者,是 caspase 家族成員中的 caspase 2、8、9、10,所扮演的角色,是將下 游的 caspase 轉為活化型,並具有蛋白質分解之能力的 caspase 3、6、 7 刺激 apoptosis 進行[62]。 其中 Caspase 3 在細胞進行 apoptosis 的過 程中是相當重要的,它扮演了細胞死亡過程中的劊子手。caspase 3 是 cysteine proteases family 中重要的一員,它將細胞中的許多蛋白質在 特定的 amino acid 上切一刀,使蛋白質被水解並失去功能。在過去的 研究中顯示,當兩個 caspase 3 被拉近以形成 dimer 時,caspase 3 將. 60.
(62) 會自行活化,並將使細胞進行不可逆轉的 apoptosis,而導致細胞的死 亡[28]。 活 化 死 亡 接 受 器 , 如 :TNFR 可 進 一 步 活 化 JNK MAPK pahtway,JNK MAPK pathway 是 MAPK 訊息傳遞路徑的其中一條, 與細胞分化及凋亡較為相關,當接受死亡接受器所傳送的訊息可以經 由調控基因或磷酸化方式進而調控 pro-apoptosis [63]。 許多證據指出,平滑肌細胞的凋亡參與粥狀動脈硬化的致病過程 而其中包括許多因素,除了 ox-LDL 外也有訊息傳遞 MAPK pathway 的參與,如:氧化低密度脂蛋白上的 Lysophosphatidylcholine 會經由 scavenger receptors 活化 p38 MAPK 而增加 monocytes CXCR2 表 現,導致人類內皮細胞的凋亡[64]。在冠狀動脈平滑肌細胞活性氧衍 生系統(Reactive oxygen-generating system)活化 Nuclear factor-?B (NF-?B)和釋放 Tumor Necrosis Factor-a(TNF-a)[65]。脂質產生 過氧化產生的 2,4-decadiena 也會對平滑肌細胞導致細胞毒性[21]。 Hsieh, et al. 2001 活性氧(ROS)則可藉由活化 caspase 進而導致平 滑肌細胞凋亡[22]。而這些結果顯示,粥狀動脈硬化過程中,血管平 滑肌細胞功能改變對於疾病嚴重度的之影響有極大關聯。. 61.
數據
+6



相關文件
附表 1-1:高低壓電力設備維護檢查表 附表 1-2:高低壓電力設備維護檢查表 附表 1-3:高低壓電力設備(1/13) 附表 2:發電機檢查紀錄表. 附表
Quality kindergarten education should be aligned with primary and secondary education in laying a firm foundation for the sustainable learning and growth of
[r]
[r]
肥大细胞内 Ca 2+ 增加可导致过敏介质释放, Ca 2+ 进入 胞浆也可导致支气管平滑肌收缩. 钙通道阻滞剂 ——抑制Ca
除了新聞報導,可以查看網路 評價(如 Google
Windows 95 後的「命令提示字 元」就是執行 MS-DOS 指令的應用
定期更新作業系統 定期更新作業系統,修 正系統漏洞,避免受到
