國立臺灣大學生命科學院生化科技學系 碩士論文
Department of Biochemical Science and Technology College of Life Science
National Taiwan University Master Thesis
利用農桿菌媒介重複轉形法提升美白菇異源蛋白質表 現量
Enhancement of protein expression in Hypsizygus marumoreus by multiple Agrobacterium
tumefaciens-mediated transformation
程致瑞 Chih-Jui Cheng
指導教授:黃慶璨 博士 Advisor: Ching-Tsan Huang, Ph.D.
中華民國 101 年 7 月
July, 2012
謝誌
首先最要感謝的是指導教授黃慶璨老師,教導我們許多做事與學習上該有的 態度以及思考方式,讓我在碩士班不光是學到如何規劃實驗,也學到如何安排以 及面對未來的人生;同時也感謝許瑞祥老師指導,使我們學習了許多菇蕈方面的 專業知識,並且給予我們不同的思考方向與觀念。口試期間承蒙生化科技系楊健 志老師、李昆達老師以及中研院常怡雍老師,在論文內容的細心審閱與口試時的 觀念引導,給予我論文寫作上很大的幫助,也讓本論文更臻完善,特此感謝!
這兩年來也感謝實驗室大家的照顧,先要謝謝王董、琬婷、玉儒、林琤、嗣 民與明玥等學長姐的指導,其中特別感謝實驗室一姊玉儒的領導,讓大家可以專 心地進行實驗,也要感謝大我一屆的瑞芸、巧鈴、盈潔、翊綸和佳麟還有信豪在 我碩一時給予的照顧;接著要感謝我的同學:友邦與浩業與我一起渡過許多間熬 的日子,與你們相處的日子很有趣,彼此之間的競爭也讓我們更加的成長;然後 要感謝承鴻、莉欣、寧瑄與家丞等學弟妹帶來了許多的趣味,讓我碩二兵荒馬亂 的生活仍可以感受到許多快樂。
此外也要感謝大學的朋友們跟我一起吃飯聊天談實驗,偶爾一次聚會是我放 鬆且享受的時光,感謝我的高中同學總是給我幫助與鼓勵,最後要感謝家人給我 的支持,讓我能夠無後顧之憂地學習並取得碩士學位。再次謝謝大家的照顧,因 為有你們,我的人生才能夠豐富與完整,祝福各位未來都能達成自己的理想。
致瑞 謹誌
中文摘要
分子農場(molecular farming)泛指利用轉基因植物生產具有高價值的重組蛋白 質,如醫藥用蛋白質。菇類於分子農場之應用近年來也受到重視,建立完善之菇 類異源表達系統將有助於分子農場的發展。過去研究已成功應用於多種菇類進行 異源基因表現。農桿菌媒介轉形法(Agrobacterium tumefaciens-mediated
transformation, ATMT)具有外源基因穩定性高的優點,然而缺點是異源蛋白質表現 量偏低。本研究使用美白菇(Hypsizygus marumoreus)為表達宿主,進行農桿菌媒介 重複轉形,期望能提升轉形株異源蛋白質表現量。本研究先以帶有萎銹靈抗性基 因(carboxin resistance gene , cbxr)為篩選標記與綠色螢光蛋白質基因(enhanced green fluorescent protein , egfp)為報導基因的農桿菌進行第一次轉形,接著再對該轉 形株以相同之農桿菌進行第二次轉形以取得單一抗藥性之二次轉形株;或是使用 另一種帶有潮黴素抗性基因(hygromycin phosphotransferase, hph)為篩選標記與綠 色螢光蛋白質基因的農桿菌進行第二次轉形,以取得雙重抗藥性之二次轉形株,
篩選過後比較一次轉形株母體與二次轉形株的綠色螢光蛋白質表現量。結果顯示 單一抗藥性之二次轉形株其綠色螢光蛋白質最高為每克總可溶性蛋白質中含有 319.58 ng,百分比為 3.19*10-5,係一次轉形株母體的4.25 倍,另外雙重抗藥性之 二次轉形株其綠色螢光蛋白質最高為每克總可溶性蛋白質中含有418.83 ng,百分 比為4.18*10-5,係一次轉形株母體的5.5 倍,顯示農桿菌媒介重複轉形法可以提升 異源蛋白質表現量。
關鍵字 : 分子農場,農桿菌媒介轉形法、美白菇、重複轉形
Abstract
Mushroom molecular farming recently attracts tremendous attention because of its application pontentials and the advantages over plant molecular farming. Agrobacterium tumefaciens-mediated transformation (ATMT) is commonly used in mushroom
transformation but its application was limited due to the low heterologous gene expression. In this study, Hypsizygus marumoreus was chosen as the expression host and was transformed by multiple ATMT in order to enhance heterologous protein expression. For the first step of multiple ATMT, A. tumefaciens harboring p0390-Cbx-Hiegfp, a Ti-plasmid contains carboxin resistance gene (cbxr) and enhanced green fluorescent protein (egfp), was used for transformation. The single transformants were re-transformed by A. tumefaciens harboring p0390-AH-Aiegfp, which contains hygromycin phosphotransferase (hph) and egfp, or re-transformed by A. tumefaciens harboring p0390-Cbx-Hiegfp. Finally, EGFP was analyzed by ELISA and the copy number of egfp was determined by real-time PCR. This study demonstrated that the heterologous gene expression was enhanced by multiple ATMT in H. marumoreus. The highest EGFP production in two-vector double transformants was 418.83 ng/g TSP (total soluble protein), which increased up to 5.5 folds in comparison with its parental single transformant. In one-vector double transformants, the highest EGFP production was 319.58 ng/g TSP, which raised to 4.25 folds and the transgene copy number accordingly increased, too. Multiple ATMT described in this study provides a new approach in improvement of the heterologous gene expression.
Key words: molecular farming, Agrobacterium tumefaciens-mediated transformation, Hypsizygus marumoreus, multiple transformation
目錄
謝誌 ... i
中文摘要 ... ii
Abstract ... iii
目錄 ... iv
表目錄 ... vi
圖目錄 ... vii
第一章 前言 ... 1
一、基因工程與異源表達 ... 1
1. 基因工程 ... 1
2. 異源表達系統 ... 1
二、分子農場 ... 4
1. 重組蛋白質生產 ... 4
2. 食用疫苗 ... 4
3. 發展現況與面臨問題 ... 5
三、食用菇類分子農場 ... 6
1. 菇類簡介 ... 6
2. 食用菇類分子農場 ... 6
四、美白菇 ... 8
1. 起源 ... 8
2. 栽培優勢 ... 8
3. 營養價值 ... 8
4. 藥理潛力 ... 9
五、食用菇類異源表達系統 ... 10
1. 表現載體 ... 10
2. 轉形策略 ... 12
六、農桿菌轉形機制 ... 14
1. Ti 質體簡介 ... 14
2. 毒性蛋白質活化 ... 14
3. T-DNA 傳送 ... 14
七、研究動機與目的 ... 16
第二章 材料與方法 ... 17
一、實驗材料 ... 17
1. 實驗菌株 ... 17
2. 質體 ... 17
3. 引子 ... 18
二、實驗方法 ... 19
1. 表現載體之建構 ... 19
2. 農桿菌媒介轉形法 ... 20
3. 農桿菌媒介重複轉形法 ... 21
4. 轉形株分析 ... 22
第三章 結果 ... 26
一、表現載體建構 ... 26
1. 重組質體確認 ... 26
二、農桿菌媒介轉形法 ... 26
1. 表現載體轉入農桿菌 ... 26
2. 美白菇與農桿菌共培養 ... 26
三、農桿菌媒介重複轉形法 ... 27
1. 二次轉形株篩選 ... 27
四、轉形株分析 ... 27
1. 轉形株 DNA 分析 ... 27
2. 轉形株蛋白質分析 ... 28
第四章 討論 ... 29
一、轉形株表現量差異 ... 29
二、轉形株純度 ... 30
三、農桿菌媒介轉形法探討 ... 30
四、重複轉形法探討 ... 31
第五章 結論與未來展望 ... 33
第六章 圖表 ... 34
第七章 參考文獻 ... 64
表目錄
表一 異源表達系統之比較 ... 34
表二 植物生產之醫藥用蛋白質發展現況 ... 35
表三 本研究使用之引子 ... 38
表四 PCR 反應條件 ... 39
表五 共培養培養基配置法 ... 40
表六 CTAB buffer 配置法 ... 40
表七 酵素免疫分析法試劑配置法 ... 41
表八 SDS 聚丙醯胺膠體試劑配置法 ... 42
表九 轉形株之綠色螢光蛋白質定量與外源基因拷貝數測定 ... 43
圖目錄
圖一 擔子菌生活史 ... 44
圖二 真菌於農桿菌媒介轉形之操作模式 ... 45
圖三 農桿菌轉形的過程 ... 46
圖四 論文架構圖 ... 47
圖五 質體圖譜 ... 48
圖六 質體 p0390-Cbx-Hiegfp 建構流程圖 ... 50
圖七 以限制酶截切確認質體 p0390-Cbx-Hiegfp 之大小 ... 51
圖八 質體 p0390-Cbx-Hiegfp 定序結果 ... 52
圖九 農桿菌 GV3101 菌落聚合酶連鎖反應 ... 54
圖十 農桿菌 LBA4404 菌落聚合酶連鎖反應 ... 55
圖十一 農桿菌轉形株與美白菇共培養過程 ... 56
圖十二 p0390-Cbx-Hiegfp 轉形株篩選過程 ... 56
圖十三 p0390-AH-Aiegfp 轉形株篩選過程 ... 57
圖十四 轉形株繼代後生長情形 ... 57
圖十五 農桿菌媒介重複轉形法的二次轉形株篩選 ... 58
圖十六 染色體 DNA 之聚合酶連鎖反應檢測 ... 59
圖十七 二次轉形株染色體 DNA 之聚合酶連鎖反應檢測 ... 59
圖十八 螢光顯微鏡觀察 ... 60
圖十九 轉形株綠色螢光蛋白質定量 ... 61
圖二十 美白菇轉形株之西方墨點法分析 ... 62
圖二十一 即時定量聚合酶鏈鎖反應 ... 63
第一章 前言
一 、基因工程與異源表達 1. 基因工程
基因工程(genetic engineering),又稱做遺傳工程或是基因改造,最早於 1973 年由Herbert Boyer 與 Stanley N. Cohen 製造出重組大腸桿菌(Escherichia coli)。此 後,基因工程技術成為生命科學領域的關鍵技術。基因工程可分成三個階段:基
因構築、轉形與基因表達。利用DNA 重組技術在細胞外把目標基因構築在載體,
使用適當的轉形方法把構築好之載體送入表達宿主進行外源基因之表現。這種藉 由基因工程使宿主生產重組蛋白質的過程稱為異源表達,異源表達已成為現今生 技產業的重要技術
2. 異源表達系統
比起從原物種分離純化蛋白質,利用異源表達生產重組蛋白質具有大量、快 速與純化方便等優點,在生技產業或是學術研究上皆提供極大的幫助。根據表達 宿主的不同,異源表達系統可以分為原核生物表達系統或是真核生物表達系統,
選擇適合的異源表達系統是重組蛋白質生產效率的關鍵[1]。
2.1 原核生物異源表達系統
大腸桿菌是目前原核生物異源表達系統的代表,大腸桿菌具有生長週期短、
易培養、易操作以及可持續產生蛋白質而不受負回饋抑制等優點,是生產重組蛋 白質常用的表達系統,但是大腸桿菌亦有不易克服的缺點,如生產異源蛋白質時 容易有內涵體(inclusion body)的形成,尤其是在生產真核生物來源之異源蛋白質時 更容易發生此現象。內涵體通常是因為蛋白質的不正確摺疊所導致,錯誤的結構 使蛋白質變為不可溶且失去正常功能,為了克服內涵體造成的問題,有研究指出 將培養溫度降低可以減少內涵體形成的機率[2, 3],或是將目標蛋白質與高度親水 性的GST tag 作成融合蛋白質亦可減少重組蛋白質形成內涵體[4]。即使如此,在 生產複雜的真核生物來源蛋白質仍舊會有蛋白質無法正確摺疊的問題,例如目標 蛋白質具有複雜的雙硫鍵結構時大腸桿菌將有很高的機率形成內涵體。大腸桿菌
缺乏後轉譯修飾,因此無法正確進行如醣基化(glycosylation)、磷酸化
(phosphorylation)、泛素化(ubiquitination)、乙醯基化(acetylation)等轉譯後修飾作用,
進而影響重組蛋白質之正確結構或是功能[5],上述這些原因造成了原核生物異源 表達系統應用的限制。
2.2 真核生物異源表達系統
2.2.1 酵母菌
釀酒酵母(Saccharomyces cerevisiae)屬於單細胞真核生物,廣泛運用於釀酒或 是麵包製作上,屬於Generally Recognized as Safe (GRAS)的微生物,因此沒有生物 安全的疑慮。酵母菌作為異源表達系統具有安全性高、可用發酵工程進行大量培 養以及擁有真核生物後轉譯修飾等優點,而釀酒酵母作為異源表達系統可追溯到 1981 年 Hitzeman 等人利用釀酒酵母生產人類干擾素[6],近年來亦有許多利用釀酒 酵母進行異源表達的例子[7, 8]。
除了釀酒酵母之外,嗜甲醇酵母菌Pichia pastoris 也成為相當成功的表達系統,
如同釀酒酵母,P. pastoris 亦具備可大量培養、操作簡單以及可進行後轉譯修飾等 優點,此外還具備強力可誘導的AOX1 啟動子,使異源表現效率更佳[9]。
2.2.2 絲狀真菌
絲狀真菌為了代謝環境中的物質,必須將大量的酵素釋放至胞外以進行作用,
因此絲狀真菌被視為可以大量外泌重組蛋白質。目前以麴菌Aspergillus spp.及木黴 菌Trichoderma spp.兩屬最為廣泛使用,某些品系在生產同源蛋白質時甚至可以達 到每公升30 克目標蛋白質的高產量[10]。然而絲狀真菌外泌異源蛋白質的效率卻 遠不如同源蛋白質,原因為對絲狀真菌外泌機制不了解以及過度表現重組蛋白質 導致其累積在細胞內無法外泌[11],因此目前絲狀真菌異源表達系統的研究著重於 了解其外泌途徑。
2.2.3 哺乳類細胞
哺乳類細胞因為具有較接近人類的後轉譯修飾系統,因此常作為生產醫藥用
蛋白質,特別是抗體這類有複雜醣基化結構的蛋白質。哺乳類細胞異源表達系統 可以分成兩類:暫時性表現(transient expression)與穩定表現(stable expression)。暫 時性表現系統可稱為COS-cell-based transient expression systems (COS TES),該系 統使用COS 細胞株進行基因表現,COS 細胞株係非洲綠猴的腎臟細胞株 CV-1 並
且經過SV40 病毒基因轉化而成,此類系統會持續地複製外源基因直到細胞死亡,
因此生產重組蛋白質較為快速但無法持續生產。穩定表現系統通常使用中國倉鼠 卵巢細胞(Chinese hamster ovary, CHO),因為可以持續性地進行外源基因表現,該 系統不論是在學術研究或是生物藥品生產,皆為哺乳類細胞異源表達系統的主流 [12]。儘管使用哺乳類細胞進行重組蛋白質生產仍有些問題存在,例如成本高、所 需時間長、操作較為繁複以及有動物性病毒汙染的風險,但目前生產醫藥用蛋白 質仍舊是以哺乳類細胞異源表達系統為主。
二 、分子農場
隨著基因工程技術的進步,越來越多生物用於重組蛋白質的生產,以應付逐 日增加的需求。分子農場(molecular farming)係利用轉基因植物來生產醫藥或是工 業用途的重組蛋白質,相較於哺乳類細胞異源表達系統,具有生產成本低、無動 物性病毒汙染風險、可大規模栽種以大量生產蛋白質等優點,近年來極受重視,
其應用範圍可分為重組蛋白質生產以及食用疫苗開發。表一為各種異源表達系統 之比較。
1. 重組蛋白質生產
植物異源表達系統自1980 年代開始發展,並於 1986 年與 1989 年成功利用菸 草細胞表現人類生長激素與單株抗體[13, 14],這也是第一個成功於植物表現的醫 藥用蛋白質與抗體,證實植物可生產構造正確之動物性蛋白質。卵白素(avidin)為 植物表達系統所生產的第一個上市商品[15],顯示植物確實可作為生物反應器來大 量生產重組蛋白質。在成本方面,植物生產重組蛋白質的成本比大腸桿菌系統低 10 到 50 倍[16]。迄今,植物表達系統已經成功的生產多種蛋白質,包括血清白蛋 白、生長激素、荷爾蒙、酵素、疫苗與抗體[17],相較於動物或微生物表達系統,
植物表達系統可為生物藥品市場提供了大量、便宜且安全的高品質蛋白質。
2. 食用疫苗
傳統疫苗是將去毒性或減毒性抗原利用針劑注射引發人體之主動免疫(active immunization),以增強對該疾病的抵抗力。這類型的疫苗必須低溫保存以避免失去 效力,且並須由醫護人員進行施打,使得施打疫苗所費不貲,造成許多發展中國 家無法全面施打疫苗。為了解決傳統疫苗保存不易、專業人員不足以及成本過高 的問題,植物食用疫苗的概念因應而生。植物食用疫苗是利用轉基因作物來生產 病原體的抗原,並且將抗原保留於植物細胞內,食用該植物便可以透過黏膜系統 得到專一性的免疫反應[18, 19, 20]。植物食用疫苗省去萃取純化的步驟,可將抗原 保存於植物組織中避免失去活性,此外食用疫苗不須由醫護人員施打,加上植物 表達系統總產量優於動物與微生物表達系統,可降低生產成本以符合發展中國家 的需要。
3. 發展現況與面臨問題
植物表達系統目前已經能夠成功生產各式蛋白質,如醫藥用蛋白質或是工業 用酵素等,亦有多種醫藥用蛋白質正在進行臨床試驗,表二為分子農場生產之醫 藥用蛋白質發展現況。
分子農場雖然倍受期待,但也面臨許多的問題,目前主要的疑慮是對生態的 危害以及食用後的安全性。美國安萬特(Aventis)公司在 1997 年將旗下的基因改造 玉米申請註冊,命名為星聯玉米(StarLinkTM),該玉米本來是做為動物飼料,不得
用於人類食品。然而2000 年時在玉米製食品之中檢測出星聯玉米的存在,使得安
萬特公司撤銷星聯玉米的註冊,也與美國與17 個州達成賠償協議[21]。除了星聯
玉米之外,近年仍有基改作物汙染的例子,例如2006 年美國生產的長粒米檢測出
基因改造稻米LLRICE 601 之存在並重創了美國稻米產業[22],以上這些例子都顯 示了目前基改作物的管理仍有很大的改善空間。基因汙染也是大眾所擔心的問題,
基因改造作物的花粉或是種子可藉由風力或昆蟲等媒介散播至環境中,並可能會 對生態系統造成影響。以上這些因素都限制了分子農場的發展,制訂適當的基改 作物管理辦法也將是發展分子農場的重點之一。
三 、食用菇類分子農場 1. 菇類簡介
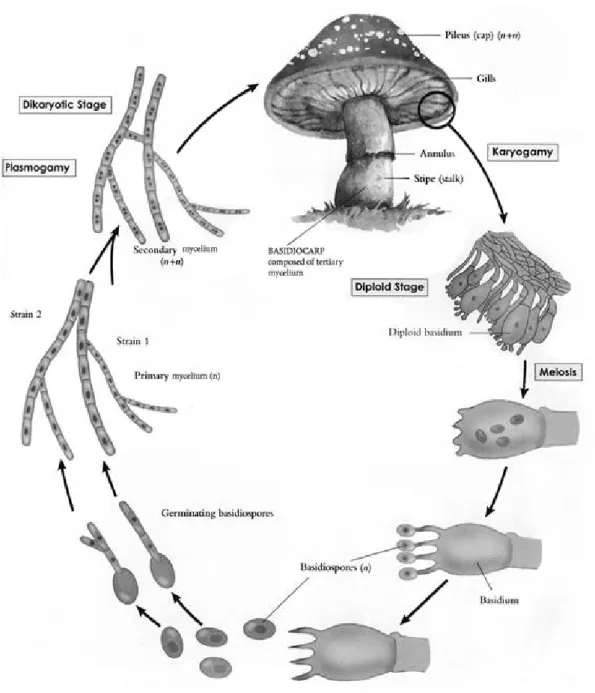
菇類或稱蕈類,原先泛指傘菌類或牛肝菌類的子實體,現今菇類係指絲狀真 菌在有性世代時形成之子實體,其大小為肉眼可見,具有菌傘(cap)、菌摺(gills)與 菌柄(stem)等構造。分類上大部分的菇類是屬於擔子菌門(Basidiomycota), 例如:
洋菇(Agaricus bisporus)、香菇(Lentinula edodes)等,少部分菇類是屬於子囊菌門 (Ascomycota),例如:羊肚菌(Morchella esculenta)等。擔子菌在有性時期會經由減 數分裂產生孢子外生於擔子柄,子囊菌是有性時期經減數分裂產生之孢子著生於 子囊內。圖一為擔子菌之生活史。
食用菇類具有鮮美之風味、高營養價值與保健功效,自古以來便廣受人們喜 愛。目前可食用的菇類有3000 多種,其中 15 種菇類已達工業化生產規模,如洋 菇、金針菇等。在營養價值方面,新鮮菇類含有約90%之水分以及 10%的乾物質,
乾物質中有27 ~ 48%的蛋白質,少於 60%的碳水化合物及 2% ~ 8%的脂肪,並含 人體所需的九種必需胺基酸、維生素B1、B2、C 與維生素 D 前驅物麥角固醇 (ergosterol)以及微量元素等營養素,是良好的營養來源。近年來研究更發現菇類含 有多種生理活性成分,如多醣體(polysaccharides)、萜類(terpenes)與免疫調節蛋白 質(immunomodulatory proteins),可提升免疫力對抗腫瘤、抗發炎、降血糖血脂[23, 24, 25, 26, 27]。
2. 食用菇類分子農場
食用菇類在分子農場的應用具有許多優勢,第一個優點是部份食用菇類可進 行工業化生產,過程皆於室內生產,可大幅減少基因汙染的風險,而且食用菇類 栽培多是利用太空包進行培養,使用完之培養基質可進行滅菌避免把基因傳遞給 土壤微生物;第二個優點是於環控培養,所以食用菇類栽培不受氣候或是季節影 響,一年四季皆可生產,加上可以立體栽種,能在有限之空間達到最大的生產效
率與生產量;第三個優點是食用菇類屬GRAS 等級,因此做為表達宿主不會有生
物安全的問題。
菇類基因轉形研究在近二十年間持續地發展,1986 年與 1987 年分別有裂褶菌 (Schizophyllum commune)及灰蓋鬼傘(Coprinus cinereus) tryptophan 營養需求菌株之
轉形研究[28, 29],上述兩種皆是擔子菌研究常用的模式物種。而食用菇類的轉形 系統則於1996 年,由洋菇表現 hygromycin B 抗性基因[30],以及使用鮑魚菇 (Pleurotus ostreatus)表現 β-glucuronidase (GUS)與 bialaphos 抗性基因[31],後續也 有許多菇類轉形的研究。現階段研究多著重在各種菇類表現報導基因或是不同轉 形策略的研究,但食用菇類想要實際應用在分子農場還有許多努力的空間。若食 用菇類異源表達系統發展成熟,相信對生技醫藥、食品與工業都會有莫大的幫助。
四 、美白菇 1. 起源
美白菇(Hypsizygus marumoreus)是由日商 Hokuto Sangyo Corporation 所研發之 食用菇類,係利用紫外光照射造成鴻喜菇(Hypsizygus marmoreus)變異後再進行交 配,最後取得白色、不具苦味的鴻喜菇白色變種,在日本稱作Bunapi-shimeji,在 台灣多稱作美白菇或雪白菇。在分子鑑定上亦確定美白菇與鴻喜菇屬於同一物種,
僅外觀顏色與味道有所差異[32]。
鴻喜菇其科學分類為真菌界(Mycota),擔子菌門(Basidiomycota),層菌綱 (Hymenomycetes),同擔子菌亞綱(Homobasidiomycetes),傘菌目(Agaricales),口蘑 科(Tricholomataceae)、玉蕈屬(Hypsizygus)。鴻喜菇之學名為 H. marmoreus,目前 確認與Hypsizygus tessulatus 是同物異名[33],此外鴻喜菇有許多中文別名,如真姬 菇、玉蕈、蟹味菇、海鮮菇、榆菇等。
2. 栽培優勢
美白菇的栽培條件與鴻喜菇相同,皆可於工廠環控進行大規模瓶栽,從接種 到採收大約需兩個月到三個月。日商Hokuto Sangyo Corporation 之菇類栽培廠估計 每間工廠年產量約為3000 公噸,該公司於台灣設立之栽培工廠年產量也可達 1100 公噸。生產美白菇子實體的溫度為14oC,相較於金針菇子實體生產最適之溫度 8 oC,
美白菇大規模栽培的能源成本較低。在培養基方面,美白菇子實體培養多使用米 糠或是玉米芯等農業廢棄物作為基質,菇類培養後所剩餘的培養基亦可作為肥料,
使資源能夠再次利用。
3. 營養價值
根據日本食品分析中心之調查,美白菇每一百克鮮重之中具有90.7 克的水、
2.4 克蛋白質、0.5 克脂質、3 克醣類、2.6 克膳食纖維與 0.8 克灰分。此外美白菇 具有高量的硒,每一百克乾重中具有21 微克的硒,是大豆的 2.9 倍,玉米的 7.5 倍,硒為人體重要微量元素,對維持人體正常生理功能極為重要,如硒為麩胱甘 肽過氧化酶(glutathione peroxidase)的重要成分,硒也跟前列腺素的合成有關,研究 亦發現硒具有抗腫瘤與抗氧化的功能[34]。
4. 藥理潛力
菇類多醣體被認為具有抗腫瘤的能力,研究發現鴻喜菇具有抗癌之潛力。鴻 喜菇多醣體中的β-(1-3)-glucan 在小鼠實驗證實具有抑制 Sarcoma 180 腫瘤生長的 功效[35],此外小鼠先餵食含有鴻喜菇粉末的飼料,再注射 20-methylcholanthrene 進行癌症誘導,發現鴻喜菇具有預防腫瘤的功效[36]。
鴻喜菇含有名為Hypsin 的蛋白質,Hypsin 為一種 ribosome-inactivating protein,
具有抑制小鼠及人類白血病細胞(leukemia cell)的能力,該蛋白質即使於 100oC 沸 水加熱10 分鐘仍能維持 60%的抑癌效果[37]。癌細胞對內皮細胞的黏附作用與癌 症轉移有密切相關,鴻喜菇中名為HM-23 的 type IV collagen-binding protein 在細 胞實驗可抑制肺癌細胞(Lewis lung carcinoma cell)之黏附作用[38]。鴻喜菇中名為 Marmorin 的 ribosome inactivating proteins (RIPs)也已知可抑制肝癌細胞株 HepG2 和乳癌株MCF-7 的增生[39]。
鴻喜菇萃取物分離出的類固醇化合物有抗結核菌感染功效,並且能夠抑制 Epstein-Barr virus (EBV)早期病毒抗原活化[40]。鴻喜菇甲醇萃取物,hypsiziprenol A9則可避免肝癌細胞HepG2 中 retinoblastoma protein (pRb)的磷酸化,使細胞停留 在G1 階段因而無法繼續生長[41]。
五 、食用菇類異源表達系統 1. 表現載體
表現載體是承載外源基因進入表達宿主的質體,表現載體不只影響外源基因 表現與否,更與基因表現量息息相關,因此建構出良好的表現載體是異源表達的 重要課題之一。
1.1 啟動子
表現載體中需要適當之啟動子以驅動下游外源基因的表現,而啟動子會影響 外源基因表現量強弱。許多異源表達之研究使用甘油醛-3-磷酸脫氫酶
(glyceraldehyde-3-phosphate dehydrogenase,gpd)基因的啟動子來驅動外源基因表現,
例如在酵母菌或絲狀真菌[42, 43, 44]。甘油醛-3-磷酸脫氫酶是生物醣解作用 (glycolysis)以及醣質新生作用(gluconeogenesis)中的重要酵素,在真核生物表現量 高,例如釀酒酵母菌的甘油醛-3-磷酸脫氫酶 mRNA 表現量佔細胞中 poly (A)+ RNA 的1~5% [45],顯示甘油醛-3-磷酸脫氫酶啟動子能夠大量地驅動下游基因的表現,
加上該啟動子有持續性驅動基因表現的特性,因此被廣泛地使用於異源表達。
現今菇類異源表達研究多使用甘油醛-3-磷酸脫氫酶啟動子進行基因表現,其 中洋菇的甘油醛-3-磷酸脫氫酶啟動子被發現可以運用於多種擔子菌,例如洋菇、
金針菇、香菇、鮑魚菇、灰蓋鬼傘與乳牛肝菌(Suillus bovinus)等等[46, 47, 48],因 此若要構築可跨物種使用的表現載體,洋菇甘油醛-3-磷酸脫氫酶啟動子是一個好 的選擇。
1.2 報導基因
報導基因(reporter gene)意指容易偵測或定量的蛋白質,藉由報導基因可輕易 地偵測啟動子強弱或宿主進行異源表現的能力。目前研究常用的有綠色螢光蛋白 質基因(green fluorescent protein gene, gfp)、β-葡萄糖苷酶基因 (β-glucuronidase gene, gusA)、冷光酶基因(luciferase gene, luc)等,其中的綠色螢光蛋白質為科學研究帶來 很大的進步。綠色螢光蛋白質吸收波長為395 與 475 nm,最大散發波長是 508 nm,
利用特定波長之光激發即可產生綠色螢光,不需外加基質[49],綠色螢光蛋白質的 用途廣泛,可以用於轉形株篩選,測試基因表現強弱,與目標蛋白質融合後進行
定位或是利用螢光共振能量轉移(fluorescence resonance energy transfer, FRET)觀察 蛋白質間的交互作用。若將綠色螢光蛋白質基因序列修改,可以得到亮度更高的 增強型綠色螢光蛋白質(enhanced green fluorescent protein gene, egfp),提升偵測的 靈敏度[50]。
1.3 篩選標記
基因工程須使用篩選標記來區分轉形株與非轉形株,其中一類篩選方法是使 用營養缺陷株(auxotroph)作為表達宿主。營養缺陷株無法於基本培養基(minimal medium)生長,藉由回補特定基因使表達宿主生產特定營養物質而存活,如此便可 篩選出轉形株,此類方法稱作營養缺陷標記(auxotrophic marker)。然而營養缺陷株 取得不易,使用營養缺陷標記必須對該表達宿主之生理生化有完整的了解。因此 目前研究多使用另一類篩選方法,利用抗生素抗性基因(antibiotic-resistant marker) 作為篩選標記,將抗生素抗性基因送入表達宿主產生可把抗生素去活化的蛋白質,
可在含有抗生素之環境下生長的便是轉形株。
在菇類異源表達研究多使用潮黴素(hygromycin B)作為篩選標記,潮黴素可抑 制原核生物與真核生物生長,其機制為妨礙核糖體之轉位作用(translocation)以及造 成錯誤轉譯(mistranslation)而影響生物體之蛋白質合成[51, 52]。潮黴素抗藥性基因 (hygromycin B phosphotransferase,hph)會對潮黴素進行磷酸化修飾導致其失去功用,
使轉形株得以在含有潮黴素之環境生長。除了潮黴素,萎銹靈(carboxin)抗性基因 也可作為菇類異源表達研究的篩選標記。萎銹靈是一種除真菌劑,對擔子菌有很 好的抑制效果,其機制為阻止電子傳遞鏈中琥珀酸去氫酶(succinate dehydrogenase) 和輔酶Q (ubiquinone)之間的電子傳遞,使得電子傳遞鏈無法進行。若是對琥珀酸 去氫酶中的鐵硫蛋白質次單元(iron-sulfur protein subunit complex II)特定序列進行 單一胺基酸突變,便可抵抗萎銹靈之作用[53]。目前已有許多菇類研究使用萎銹靈 抗性基因作為篩選標記,例如:香菇、灰蓋鬼傘、鮑魚菇與美白菇等[54, 55, 56, 57]。
使用萎銹靈抗藥性基因作為篩選標記的好處是此抗性基因屬於同源性基因,可避 免在發展菇類食用疫苗時造成人體過敏現象。
1.4 分生策略
適當的分生策略可有效提升異源表達的表現量,包括於目標基因前加上內含 子(intron)或是於目標基因後端加上內質網停留訊息胜肽(endoplasmic reticulum retention signal peptides)。研究發現於目標基因前端添加內含子可以增加 mRNA 的 累積,於洋菇、灰蓋鬼傘、裂褶菌及白腐型真菌(Phanerochaete chysosporium)的研 究中皆可發現內含子之有無確實影響了報導基因的表現[58, 59, 60]。
在目標基因後端加上內質網停留訊號可使目標蛋白質轉譯後停留在內質網 (endoplasmic reticulum, ER)或高基氏體(Golgi body)中,使目標蛋白質完整地摺疊與 進行修飾,進而提升目標蛋白質於細胞中的累積量[61]。
2. 轉形策略
基因轉形方法可分為兩類:非載體媒介轉形法(vector-free gene transfer)或是載 體媒介轉形法(vector-mediated gene transfer)。常用於菇類轉形研究的非載體媒介轉 形法包括電穿孔法(electroporation)、Polyethylene glycol (PEG)媒介轉形法與限制酶 媒介插入法(restriction enzyme-mediated integration, REMI)法;常用於菇類轉形研究 的載體媒介轉形法則是農桿菌媒介轉形法(Agrobacterium tumefaciens-mediated transformation)。
2.1 電穿孔法
電穿孔的原理是使用瞬間高電場使細胞膜形成孔洞,外源基因經由孔洞而進 入表達宿主,當電場消失後細胞膜可回復,使外源基因停留在細胞內。不同的細 胞在細胞膜穿孔所需之電場閾值不同,利用電穿孔法必須先測試最適電穿孔條件。
電穿孔適用於大部分的細胞,許多物種如細菌、絲狀真菌、植物及動物皆有利用 電穿孔轉形成功的例子,然而電穿孔法在菇類轉形的缺點是外源基因穩定性不 高。
2.2 PEG 媒介轉形法
PEG 是極具親水力的分子,高濃度 PEG 可聚集原生質體(protoplast)並使之融 合,亦可促使細胞產生胞飲作用攝入DNA。大多數 PEG 媒介轉形法需要以原生質 體為材料,但菇類的原生質體製備過程繁複且不易再生,使得轉形率偏低,同時
也有外源基因穩定性不高的缺點,使得PEG 媒介轉形法在菇類轉形發展受到限 制。
2.3 限制酶媒介插入法
限制酶媒介插入法的操作與PEG 媒介轉形法類似,但多了限制酶酵素處理,
限制酶酵素可進入原生質體的細胞核,對表達宿主染色體進行截切,宿主在修復
染色體的過程便有機會與表現載體進行黏合。與PEG 媒介轉形法相比,限制酶媒
介插入法可提升轉形率10 倍之多,同時也具有較佳的外源基因穩定性[62]。
2.4 農桿菌媒介轉形法
腫瘤農桿菌(A. tumefaciens)是一種存在於土壤中的革蘭氏陰性菌(Gram
negative),可藉由傷口感染植物,促使植物形成冠狀腫瘤(crown gall tumor)。腫瘤 農桿菌可將自身基因嵌入受感染宿主染色體,利用腫瘤農桿菌將欲表現之基因送 入表達宿主,稱作農桿菌媒介轉形法。早期農桿菌媒介轉形法僅應用於植物,但 近年來發現酵母菌、絲狀真菌甚至哺乳類細胞都可以藉由農桿菌媒介轉形法進行 基因表達[63, 64, 65]。目前已有許多農桿菌媒介轉形法成功應用於菇類轉形之例子,
相較於其他的轉形方法,農桿菌媒介轉形法之優點為不需製備原生質體,操作簡 單、轉形率以及外源基因穩定性高,顯示農桿菌媒介轉形法於菇類轉形系統研究 深具發展潛力。圖二為真菌於農桿菌媒介轉形之操作模式。
然而以農桿菌媒介轉形法應用於菇類異源基因表現時面臨之問題是異源蛋白 質表現量不高,以金針菇為表達宿主的異源表達研究顯示綠色螢光蛋白質含量佔 全部可溶性蛋白質(total soluble protein, TSP)的比例低於 0.01% [66]。可能是因為農 桿菌的T-DNA 大多是以單一拷貝數(copy number)隨機插入,低基因拷貝數使得蛋
白質表現量偏低。目前菇類轉形研究顯示T-DNA 插入拷貝數介於一至四個之間,
但以單一拷貝數居多[67],如何提升異源蛋白質表現量是目前菇類轉形系統重要的 課題。
六 、農桿菌轉形機制 1. Ti 質體簡介
腫瘤農桿菌其致病過程主要由腫瘤引發質體(Tumor-inducing plasmid, Ti
plasmid)所調控。腫瘤引發質體為一環狀雙股 DNA 分子,可概分為:Transferred DNA (T-DNA)區域以及毒性蛋白質基因群(vir genes)。T-DNA 區域由兩段約 25 bp 的序 列Right border (RB)與 Left border (LB)所夾帶,該序列會於農桿菌感染宿主時送入
宿主染色體,原生型的腫瘤引發質體其T-DNA 區域內具有可誘使植物宿主產生激
素的基因,進而造成異常分裂增生的腫瘤[68, 69]。
2. 毒性蛋白質活化
圖三為農桿菌感染植物細胞之過程。植物受傷部位釋出的胺基酸、有機酸與 醣類吸引農桿菌附著,同時植物受傷部位釋出的酚類化合物活化農桿菌VirA/VirG。
VirA/VirG 為二元調節系統(two-component signal-transduction system),當 VirA 與酚 類化合物結合會促使VirA 發生自體磷酸化(autophosphorylation),並進一步促使 VirG 發生磷酸化,磷酸化的 VirG 將作為轉錄活化子(transcriptional activator)調控其 他毒性蛋白質基因之表現。毒性蛋白質表現後,具有內切酶(endonuclease)活性的 VirD1 及 VirD2 辨認 RB 與 LB 序列並切下 T-DNA,被切下的 T-DNA 則與 VirD2 形成未成熟T-DNA 複合體(immature T-complex),以避免被分解[70]。
3. T-DNA 傳送
農桿菌VirB/D4 複合體於自身與受感染細胞之間形成第四型分泌系統(Type IV secretion system, T4SS)以傳送 T-DNA,形成之第四型分泌系統可將未成熟 T-DNA 複合體與 VirE2 以及 VirF 等毒性蛋白質一併送入宿主細胞,在宿主細胞中 未成熟T-DNA 複合體與 VirE2 結合成為成熟 T-DNA 複合體[68, 71]。
4. T-DNA 嵌入
T-DNA 複合體上的 VirD2 與 VirE2 都有核酸定位訊號(nuclear localization signal, NLS)序列,當 T-DNA 複合體進入宿主細胞後,VirD2 與 VirE2 可分別藉由 轉運蛋白質(importin-α family)以及 VirE2 交互作用蛋白質 1 及 2 (VirE2 interacting
protein 1, VIP1 and 2, VIP2)的幫助,促使 T-DNA 進入細胞核[72]。農桿菌 T-DNA 進入細胞核後,將以隨機的方式嵌入宿主染色體,嵌入的詳細機制至今尚無定論,
推測是於染色體雙股螺旋斷裂(Double-strand break)之處嵌入。在釀酒酵母的農桿菌 媒介轉形研究,發現T-DNA 可藉由同源重組(homologous recombination, HR)或是 非同源末端接合(nonhomologous end joining, NHEJ)嵌入,並且發現嵌入的模式與宿 主的因子有密切相關。若將參與同源重組的因子剔除,酵母菌會使用非同源末端 接合嵌入T-DNA,反之亦然[63],而在植物的系統 T-DNA 主要是以非同源性末端 接合進行嵌入。目前推測T-DNA 複合體會利用宿主本身的蛋白質幫助 T-DNA 嵌 入,T-DNA 複合體中的 VirD2 與 VirE2 功用可能是吸引相關蛋白質至雙股螺旋斷 裂處參與反應[73]。
七 、研究動機與目的
菇類轉形系統的開發與食用菇類分子農場的發展息息相關,本實驗室先前已 建立菇類電穿孔轉形系統,然而電穿孔法基因穩定性低,不利於菇類異源表達系 統的應用。為改善基因穩定性的問題,本實驗室轉而發展菇類農桿菌媒介轉形法,
並於金針菇、美白菇以及鮑魚菇成功地進行異源表達,但也面臨異源蛋白質表現 量不高的問題。本研究將利用美白菇之農桿菌媒介轉形系統進行重複轉形,藉由 多次轉形以增加送入宿主的外源基因拷貝數,進而提高異源蛋白質表現量。為達 成此目的所需完成的目標包括:
1. 建構美白菇異源表現載體
建構具有美白菇甘油醛-3-磷酸脫氫酶(gpd)啟動子、萎銹靈抗藥性基因以及報 導基因egfp 的表現載體。
2. 進行農桿菌媒介重複轉形法 2.1 將表現載體轉入農桿菌株。
2.2 以美白菇菌絲塊作為轉形材料進行農桿菌媒介轉形。
2.3 篩選轉形株。
2.4 對轉形株再次進行農桿菌媒介轉形。
3. 轉形株分析
3.1 利用聚合酶連鎖反應(PCR)確認外源基因插入與否。
3.2 利用西方墨點法(western blotting)和酵素免疫分析法(enzyme-linked immunoassay, ELISA)進行蛋白質定性與定量分析。
3.3 以即時定量聚合酶鏈鎖反應(real-time PCR)測定外源基因拷貝數。
3.4 以螢光顯微鏡觀察表現EGFP 之轉形株。
本論文的研究架構如圖四所示。
第二章 材料與方法
一 、實驗材料 1. 實驗菌株
1.1 真菌
本研究所使用之菌種為美白菇(Hypsizygus marumoreus),於台北頂好市場購入 後分離出菌絲進行培養。美白菇菌種保存於Potato dextrose agar (PDA)斜面培養基,
培養溫度為14oC。一般培養在固態 MYGPA 平板培養基(1% malt extract, 1% yeast extract, 0.5% glucose, 0.3% peptone, 1.5% agar)或 MYGP 液態培養基,培養溫度為 23oC。
1.2 細菌
大腸桿菌Escherichia coli DH5α 作為基因轉殖及序列保存之宿主細胞,一般培 養於固態平板培養基Luria-Bertani (LB)或於液態培養基 LB broth 在 37oC 下以 250 rpm 振盪培養。長期保存則使用含 15%甘油之 LB broth,於 -80oC 下保存。
農桿菌Agrobacterium tumefaciens LBA4404 和 GV-3101,作為農桿菌轉形媒介,
一般培養於固態平板培養基LB 或於液態培養基 LB broth 在 28oC,250 rpm 振盪培 養。長期保存則使用含25%甘油之 LB broth,於 -80oC 下保存。
2. 質體
本研究使用的質體是以pCAMBIA0390 為骨架進行建構或農桿菌媒介轉形法。
p0390-AH-Aiegfp、pHi-hyg-egfpS、p0390-sdi1-ORFM、為本實驗室前人[57, 66]所 建構,p0390-Cbx-Hiegfp 於本研究完成。質體詳細內容如下:
2.1 pCAMBIA-0390
pCAMBIA-0390 質體大小為 6.8 kb,主要應用於農桿菌媒介轉形系統。是一種 雙偶型載體(binary vector),帶有 NOS 3’UTR poly A 終結子、left border (LB)、right
border (RB)序列、pUC9 載體多限制酶切位及卡那黴素抗藥性基因(kanamycin resistance gene)。
2.2 pHi-hyg-egfpS
pHi-hyg-egfpS 質體大小為 7.1 kb,此質體以 yT&A 質體作為骨架進行構築, 帶 egfp 與潮黴素抗性基因(hygromycin phosphotransferase, hph),兩者皆以美白菇 gpd 啟動子啟動。
2.3 p0390-AH-Aiegfp
p0390-AH-Aiegfp 質體大小為 9.4 kb,此質體以 pCAMBIA-0390 做為骨架進 行建構,egfp 基因為報導基因以及 hph 為篩選標記,兩者皆以洋菇 gpd 啟動子啟 動。
2.4 p0390-sdi1-ORFM
p0390-sdi1-ORFM 質體大小為 8.8 kb,此質體以 pCAMBIA-0390 做為骨架進 行建構,萎銹靈抗性基因(carboxin resistance gene , cbxr)為篩選標記,此基因以美白 菇sdi1 啟動子啟動。
2.5 p0390-Cbx-Hiegfp
p0390-Cbx-Hiegfp 質體大小為 11.1 kb,此質體以 pCAMBIA-0390 做為骨架進 行建構,egfp 基因為報導基因與 cbxr為篩選標記,egfp 基因以美白菇 gpd 啟動子 啟動,萎銹靈抗性基因以美白菇sdi1 啟動子啟動。
上述質體之圖譜如圖五所示。
3. 引子
本研究所使用的引子為委託生技公司合成(基龍米克斯,台灣)。詳細序列整理於表 三。
二 、實驗方法 1. 表現載體之建構
1.1 質體抽取
將攜帶質體p0390-sdi1-ORFM 的大腸桿菌菌株於含有 50 μg/ml kanamycin 之 3 mL LB 中 37oC 隔夜培養後,使用 Miniprep plasmid extraction kit (Genemark,台灣) 抽取質體。
1.2 聚合酶鏈鎖反應
將稀釋至適當濃度之質體pHi-hyg-egfpS 作為模版,並且使用引子組 (AseI-HiGPD-F / XmaI-Nos-R)擴增出目標片段(AseI-HiGPD-egfp-NOS- XmaI),PCR 之反應物組成為0.2 μM 正向引子、0.2 μM 反向引子、0.1 μM dNTP、1.5 μL 10x Pfu buffer、0.075 μL Pfu DNA polymerase (5 U/μL)、7.225 μL ddH2O,總體積為 15 μL。
反應條件見表四。反應完之產物使用洋菜膠體進行電泳確認產物大小。
1.3 洋菜膠體電泳
將DNA 與 6x 追蹤染劑混合後,以 1%洋菜膠體進行電泳,於 0.5x TAE buffer (40 mM Tris-acetate, 1mM EDTA, pH8.0)以電壓 140 伏特進行電泳 20 分鐘。經 ethidium bromide 外染 10 ~ 15 分鐘後,在 UV box 上照相、觀察結果。
1.4 限制酶截切
將質體p0390-sdi1-ORFM 與 PCR 產物 AseI-HiGPD-egfp-NOS- XmaI 分別使用 限制酶AseI 與 XmaI 於 37oC 反應兩個小時。反應後溶液使用 DNA clean up kit (Genemark,台灣)進行純化。
1.5 質體黏合與轉形
確認純化後DNA 濃度與大小,以莫耳數比 vector:insert = 3:1 的條件,加 入1 μL T4 接合酶、10x 緩衝液 A、10 x 緩衝液 B (益生,台灣)以及無菌水使最終 體積為10 μL,於 4oC 反應隔夜。反應後溶液使用 heat-shock 轉形法將 DNA 送入
大腸桿菌DH5α 勝任細胞中,於含 50 μg/ml kanamycin 之 LB 固態平板培養基上培 養16 ~ 18 小時。各質體建構流程如圖六所示。
1.6 大腸桿菌轉形株篩選
以牙籤挑取上一步驟之大腸桿菌菌落,加入專一性引子與試劑進行菌落聚合 酶連鎖反應(Colony PCR),PCR 反應條件見表四。反應完成後以洋菜膠體電泳分析,
確認目標產物大小無誤後,抽取質體進行限制酶截切確認並將正確質體進行核酸 定序(基龍米克斯,台灣)。
2. 農桿菌媒介轉形法
2.1 農桿菌勝任細胞製備
挑取農桿菌單一菌落至3 mL LB broth,在 28oC,250 rpm 下振盪培養 48 小時,
使OD600吸光值達到1.0-1.5 之後,稀釋菌液 100 倍以 100 mL LB broth 大量培養 18 小時,使 OD600吸光值達到1.5-2.0。將菌液以 4oC,3000 g 離心十分鐘,去除 上清液後以等體積的冰鎮無菌水將菌體懸浮,如此重複五次,最後一次以400 μL 冰鎮的10%無菌甘油將菌體懸浮,以每管 40 μL 分裝至微量離心管,置於 -80oC 保存。
2.2 以電穿孔法將表現載體移入農桿菌
將2 μL 已構築好之表現載體加入 40 μL 農桿菌勝任細胞,均勻混合並於冰上 放置2 分鐘,加入電穿孔石英管中,將管壁之水分以拭鏡紙擦乾,以 1.25 kV 進行 電穿孔(Chamber gap : BTX disposable cuvette P/N 610 (1 mm gap),field strength : 12.5 kV/cm,capacitance : 25 F,resistance : 200 ohms,pulse Length : 5.0 msec)。
操作完成後於無菌操作台內加入400 μL LB broth,混勻後全數吸到微量離心管中,
於室溫靜置培養2.5 小時。取 25 ~ 50 μL 菌液均勻塗抹在含有 50 μg /mL kanamycin 之LB 固態平板培養基上,以 28oC 培養兩天。
2.3 農桿菌轉形株篩選
以牙籤挑出上一步驟之農桿菌菌落,加入專一性引子與試劑進行菌落聚合酶 連鎖反應(Colony PCR),反應條件見表四。反應完成後以洋菜膠體電泳分析,確認 目標產物大小。
2.4 農桿菌培養
將農桿菌轉形株於3 mL 含 50 μg/ml kanamycin 之 LB broth,以 28oC、250 rpm 振盪培養48 小時。接著取 50 μL 菌液至 50 mL 含 50 μg/ml kanamycin 之 LB broth,
以28oC、250 rpm 振盪培養 12 ~ 16 小時。以 4oC,4,000 g 之條件離心十分鐘後將 菌體以含有200 μM 誘導物 acetosyringone (AS)之 IM broth 懸浮均勻使菌液 OD600 = 1,並準備與美白菇菌絲塊共培養。IM 培養基依表五配製而得。
2.5 真菌與農桿菌共培養
以打孔器將生長於MYGpA 之美白菇菌絲,打成數個直徑約 0.5 mm 的顆粒菌 絲塊(Modified mycelial pellets, MMP),菌絲塊加入以 IM broth 懸浮後之農桿菌菌液 並靜置培養4 ~ 6 小時,接著將菌絲塊置於含有誘導物 AS 之 IM 平板培養基上,
共培養三到六天。
2.6 農桿菌之去除
共培養完成後,以含有200 μM cefotaxime 的無菌水洗滌五次並以滅菌後的擦 手紙吸乾殘留液體,將農桿菌從菌絲塊中去除。
2.7 轉形株篩選
將農桿菌去除後之菌絲塊置於選擇性培養基進行篩選。使用含有200 μM cefotaxime 以及 7.5 μg/ml hygromycin B 的 MYGP 培養基確認表現潮黴素抗性基因 之p0390-AH-Aiegfp 轉形株;使用含有 200 μM cefotaxime 以及 3 μg/ml carboxin 的 MYGP 培養基確認表現萎銹靈抗性基因之 p0390-Cbx-Hiegfp 轉形株。以上皆於 23oC 靜置培養。
3. 農桿菌媒介重複轉形法
p0390-Cbx-Hiegfp 轉形株經過至少二次繼代後對該轉形株再次進行農桿菌媒
介轉形,即稱作農桿菌媒介重複轉形法或是重複轉形。重複轉形分為兩種策略,
一是雙質體轉形策略(two-vector transformation),另一種稱為單質體轉形策略 (one-vector transformation)。
3.1 雙質體轉形策略
雙質體轉形策略係利用帶有表現載體p0390-AH-Aiegfp 之農桿菌對 p0390-Cbx-Hiegfp 轉形株再次進行農桿菌媒介轉形,轉形後使用含有 200 μM cefotaxime、3 μg/ml carboxin 以及 3 μg/ml hygromycin B 的 MYGP 培養基進行篩選,
即可得到雙重抗藥性之二次轉形株。
3.2 單質體轉形策略
單質體轉形策略係利用帶有表現載體p0390-Cbx-Hiegfp 之農桿菌再次對 p0390-Cbx-Hiegfp 轉形株再次進行農桿菌媒介轉形法,便可取得單一抗藥性之二次 轉形株,篩選培養基為含有200 μM cefotaxime 與 3 μg/ml carboxin 的 MYGP 培養 基。
4. 轉形株分析
4.1 染色體 DNA 抽取
將美白菇轉形株接種於15 mL MYGP 液態培養基中,於 23oC 靜置培養。之後 收取液態培養的菌絲體,將之冷凍乾燥後使用液態氮研磨成粉末,20 mg 菌絲粉末 加入500 μL CTAB buffer,並以組織研磨棒研磨,研磨後加入 3 μL 2-mercaptoethanol,
混合均勻後於65oC 放置 30 分鐘。接著加入 500 μL CI (chloroform : isoamylalcohol = 24 : 1),溫和地上下旋轉混合,再以 4oC,12,000 g 的條件離心 10 分鐘。取出上清 液並加入500 μL PCI (phenol : chloroform : isoamylalcohol = 25 : 24 : 1),溫和地上下 旋轉混合,再以4oC,12,000 g 的條件離心 10 分鐘。取出上清液並加入 0.6 倍體積 之isopropanol,溫和地上下旋轉混合後於 -20oC 隔夜放置。接著以 4oC,12,000 g 的條件離心10 分鐘,倒去上清液。加入 500 μL wash buffer 輕微搖晃後靜置 2 分鐘,
接著以4oC,12,000 g 的條件離心 2 分鐘,倒去上清液並於抽氣櫃將多餘液體抽乾。
最後加入100 μL ddH2O 將其回溶。CTAB buffer 與 wash buffer 如表六配置。
4.2 聚合酶連鎖反應
將轉形株和野生株的染色體 DNA 稀釋至適當倍數,取 5 μL 當做模版,PCR 之反應物組成為0.2 μM 正向引子、0.2 μM 反向引子、0.1 μM dNTP、1.5 μL 10x Taq buffer、0.075 μL Taq DNA polymerase (5 U/μL)、7.225 μL ddH2O,總體積為 15 μL。
所用引子以及反應條件見表四。反應產物使用洋菜膠體進行電泳確認產物大小。
4.3 螢光顯微鏡觀察
利用螢光顯微鏡Eclipse E600 (Nikon, Kanagawa, Japan)觀察菌絲,以藍光(395 nm)激發下散發出綠色螢光(509 nm)之菌絲即為成功表現綠色螢光蛋白質之轉形株。
螢光視野下的相機曝光時間皆為4 秒。
4.4 蛋白質粗萃取
收取液態培養的菌絲體,將之冷凍乾燥後使用液態氮研磨成粉末,每50 mg
菌粉加入1 mL 蛋白質萃取液(300 mM NaCl, 50 mM sodium phosphate, 1 mM PMSF, 0.1% Triton X-100, pH 7.4),使用超音波機(Sonics & Materials Inc., USA)進行均質化,
於冰上放置一小時後,以4oC,13,000 g 的條件離心 30 分鐘,收取上清液,接著 再使用相同條件離心一次並收取上清液,於 -80oC 保存。
4.5 蛋白質定量
使用bicinchoninic acid (BCA)法進行可溶性總蛋白質定量。使用 bovine serum albumin (Sigma, Germany)配置 0、0.03125、0.0625、0.125、0.25、0.5、1、2 mg/ml 共八種濃度之標準品,將10 μL 標準品或待測蛋白質萃取液加至 96 孔微量滴定盤,
再加入 200 μL BCA working reagent (Pierce, Rockford, IL, Reagent A:B, 50:1, v/v)於 37oC 避光呈色 30 分鐘後偵測 562 nm 之吸光值。將標準品測得之吸光值製作蛋白 質含量校正曲線,以換算粗萃液的可溶性總蛋白質含量。
4.6 酵素免疫分析法
酵素免疫分析法(enzyme-linked immunosorbent assay, ELISA)依目的分為多種
類型,本研究使用間接型酵素免疫分析法(Indirect sandwich ELISA)定量轉形株之綠 色螢光蛋白質。先以coating buffer 將 mouse anti-GFP 單株抗體(ab1218, Abcam, UK)稀釋 6,000 倍,加 100 μL 於 ELISA 專用 96 孔微量滴定盤中,於 23oC 靜置 16 小時後以200 μL PBST buffer 清洗三次後,加入 300 μL gelatin-NET 於 37oC 下靜置 2 小時後去除多餘液體備用。以 EGFP (BioVision, USA)當作標準品,以 gelatin-NET 稀釋成6、5、4、3、2、1、0.5 以及 0 ng/ml 共八種濃度標準品,將 100 μL 標準品 溶液和轉形株粗萃取液加至96 孔微量離心盤,於 37oC 靜置 1 小時,再以 PBST 清洗四次後,加入100 μL 以 gelatin-NET 稀釋 5,000 倍之 rabbit anti-GFP 多株抗體 (ab6556, Abcam, UK)於 37oC 靜置 1 小時,以 PBST 清洗五次後,加入 100 μL 以 gelatin-NET 稀釋 5,000 倍之 goat anti-rabbit IgG HRP conjugate (PerkinElmer, USA) 於37oC 靜置 1 小時,以 PBST 清洗六次後,加入 100 μL 3,3’,5’5-tetramethylbenzidine 基質溶液(TMB One Component HRP Microwell Substrate, BioFX, USA)避光呈色 30 分鐘後,以2 N H2SO4終止反應,偵測450 nm 之吸光值。將標準品測得之吸光值 製作標準曲線,以換算轉形株粗萃液的綠色螢光蛋白質含量。酵素免疫分析法所 用之試劑配方如表七。
4.7 西方墨點法
將各轉形株之蛋白質粗萃取液加入5x SDS 樣品緩衝液混合均勻,於 99oC 加熱 10 分鐘後冷卻至室溫備用。將 12% SDS 聚丙醯胺膠體架設於電泳槽上,再把樣品 加入焦集膠體齒梳槽,先以電壓80 伏特通電至蛋白質焦集成一直線,再提高至 150
伏特分離蛋白質約1 小時後,將膠體取出浸泡於轉印緩衝液中。轉印前先將
Polyvinylidendifluroid (PVDF) membrane 浸泡於 100%甲醇中 10 分鐘,再以半乾式 電泳轉印槽(Genmedika Biotechnology,台灣),依序放置以轉印緩衝液浸濕之濾紙 兩片、電泳膠片、PVDF 膜、轉印緩衝液浸濕之濾紙兩片,使用 60 mA,30 分鐘 之條件將蛋白質轉印至PVDF 膜上。取出轉印後 PVDF 膜以 gelatin-NET 室溫振盪 浸洗1.5 小時,倒去 gelatin-NET 後加入以 gelatin-NET 稀釋 6,000 倍之 mouse anti-GFP 一次抗體(Living colors A.v. Monoclonal [JL-8], USA)室溫振盪浸洗 1 小時,
再以PBST buffer 清洗三次,每次 10 分鐘。加入以 gelatin-NET 稀釋 5,000 倍之 goat anti-mouse IgG AP conjugate 二次抗體(PerkinElmer, USA)室溫振盪浸洗 1 小時,再
以PBST buffer 清洗四次,每次 10 分鐘。最後使用 NBT/BCIP 基質液避光呈色,
直至紫色條帶出現後以清水漂洗終止反應。上述所用之試劑配方如表八。
4.8 即時定量聚合酶鏈鎖反應
利用即時定量聚合酶鏈鎖反應(real-time PCR)測定一次轉形株母體與二次轉形 株之外源基因拷貝數,目標基因為egfp,所用之引子對為 egfp362-F 與 egfp468-R,
參考基因為GAPDH,所用之引子對為 HmGPD-F 與 HmGPD-R。取 5 μl 轉形株 DNA 為模版,其餘反應物組成為0.2 μM 正向引子、0.2 μM 反向引子、4.2μl ddH2O 與 10 μl GM SYBR qPCR Mix (Genemark,台灣),總體積為 20 μl,反應條件為 94oC,
3 min 後接著進行 35 次循環的 94oC,15 sec 與 60oC,31 sec。計算方法使用比較性 Ct 法(Comparative Ct method),ΔCt 代表 egfp 之 Ct 值減去 GAPDH 之 Ct 值,ΔΔCt 代表二次轉形株的ΔCt 減去一次轉形株的 ΔCt,最後以 2-ΔΔCt表示差距倍數。
第三章 結果
一 、表現載體建構 1. 重組質體確認
1.1 p0390-Cbx-Hiegfp
將質體p0390-sdi1-ORFM 與 PCR 產物 AseI-HiGPD-egfp-NOS- XmaI 進行接合 反應並將反應後的質體送入大腸桿菌DH5α 勝任細胞,接著以引子對 egfp-F/egfp-R 進行colony PCR,挑取帶有目標質體的菌落,將質體抽出後以限制酶 BglII 與 AseI 進行截切,可得到1.2 kb、2.2 kb 以及 7.6 kb 的預期片段,如圖七。將質體
p0390-Cbx-Hiegfp 進行定序以確認序列無誤,如圖八。
二 、農桿菌媒介轉形法 1. 表現載體轉入農桿菌
將表現載體p0390-Cbx-Hiegfp 與 p0390-AH-Aiegfp 分別以電穿孔法送入農桿 菌GV3101 勝任細胞,接著以引子對 egfp-F/egfp-R 進行 colony PCR,順利轉形者 可於洋菜膠體電泳後,得到0.7 kb 的片段,如圖九。表現載體 p0390-Cbx-Hiegfp 與p0390-AH-Aiegfp 亦送入農桿菌 LBA4404 勝任細胞,並以引子對 egfp-F/egfp-R 進行colony PCR,結果如圖十。
2. 美白菇與農桿菌共培養
2.1 共培養過程
將野生型美白菇菌絲塊(Modified mycelial pellets, MMP)與帶有表現載體的農
桿菌轉形株進行共培養,於IM 平板培養基上共培養的情況如圖十一。
2.2 篩選轉形株
將共培養後的菌絲塊去除農桿菌並轉移至選擇性培養基上進行轉形株篩選。
p0390-Cbx-Hiegfp 轉形株於含有 200 μM cefotaxime 與 3 μg/ml carboxin 的選擇性培
養基進行篩選,如圖十二。p0390-AH-Aiegfp 轉形株於含有 200 μM cefotaxime 與 7.5 μg/ml hygromycin B 的選擇性培養基進行篩選,如圖十三。
上述轉形株於選擇性培養基上生長一定大小後,挑取外緣之新生菌絲轉移至 新的選擇性培養基上繼續培養,以排除偽陽性的情況,如圖十四。
三 、農桿菌媒介重複轉形法 1. 二次轉形株篩選
進行雙質體轉形策略之重複轉形後,使用同時含有3 μg/ml carboxin、3 μg/ml hygromycin B 以及 200 μM cefotaxime 之選擇性培養以篩選出二次轉形株。結果顯 示p0390-Cbx-Hiegfp 轉形株與 p0390-AH-Aiegfp 轉形株無法於該培養基上生長,
如圖十五(A)與圖十五(B)。二次轉形株則是能夠生長於雙重抗生素的選擇性培養基,
如圖十五(C)。藉由表現兩種抗生素抗性,可證實重複轉形之可行性。
四 、轉形株分析 1. 轉形株 DNA 分析
1.1 一次轉形株聚合酶連鎖反應檢測
將p0390-Cbx-Hiegfp 轉形株與 p0390-AH-Aiegfp 轉形株之染色體 DNA 抽出後,
分別以引子對egfp-F/egfp-R 以及 ApGPD-F/hph-R 進行聚合酶連鎖反應,以確認外 源基因是否送入轉形株。p0390-Cbx-Hiegfp 轉形株的檢測,如圖十六(A)所示,染 色體DNA 可以擴增出預期的 0.7 kb 片段,p0390-AH-Aiegfp 轉形株的檢測,如圖 十六(B)所示,染色體 DNA 可以擴增出預期的 1.3 kb 片段。以上結果皆顯示外源 基因順利送入轉形株。
1.2 二次轉形株聚合酶連鎖反應檢測
藉由雙質體轉形策略重複轉形所得到的二次轉形株,抽出染色體DNA 後,以
引子對ApGPD-F/hph-R 進行聚合酶連鎖反應,以確認 p0390-AH-Aiegfp 之 T-DNA 確實送入二次轉形株之中。結果如圖十七所示,染色體DNA 可以擴增出預期的 1.3 kb 片段,證實第二次轉形成功把外源基因送入宿主。
2. 轉形株蛋白質分析
2.1 螢光顯微鏡觀察
利用螢光顯微鏡觀觀察野生型美白菇菌絲與轉形株菌絲是否可以散發出綠色 螢光。如圖十八所示,在藍光激發下,轉形株可以發出綠色螢光,但是各轉形株 的綠色螢光強度普遍不強,因此難以藉由螢光顯微鏡來區別一次轉形株與二次轉 形株,僅可確認綠色螢光蛋白質之生成。
2.2 酵素免疫分析法
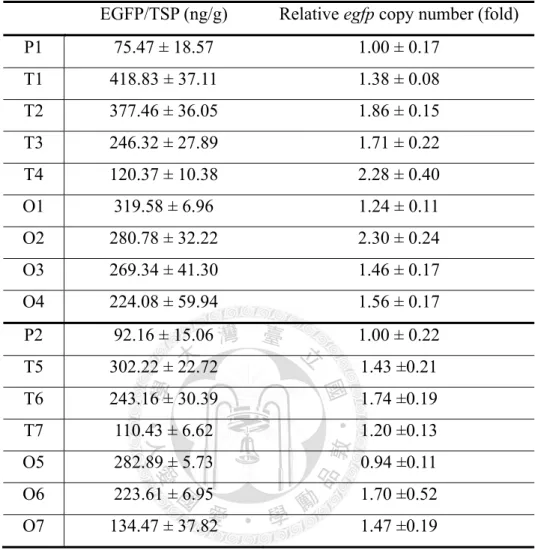
轉形株經過粗萃取法取得總可溶性蛋白質後,以ELISA 分析其綠色螢光蛋白
質含量,結果如圖十九與表九,一次轉形株母體測得綠色螢光蛋白質為每克總可 溶蛋白質含有75.47 ng,而雙質體轉形策略之二次轉形株最高可測得每克總可溶蛋 白質含有418.83 ng,是一次轉形株母體的 5.5 倍,而單質體轉形策略之二次轉形 株最高可測得每克總可溶蛋白質含有319.58 ng 的綠色螢光蛋白質,是一次轉形株 母體的4.25 倍,顯示兩種重複轉形策略皆可提升綠色螢光蛋白質的表現量。
2.3 西方墨點法
將轉形株蛋白質粗萃液進行西方墨點法分析,確認轉形株綠色螢光蛋白質大 小之正確性。結果如圖二十,蛋白質粗萃液中可測得約27 kDa 的綠色螢光蛋白質。
2.4 即時定量聚合酶鏈鎖反應
利用即時定量聚合酶鏈鎖反應相對定量測定轉形株之egfp 拷貝數,進而確認
二次轉形株之外源基因拷貝數是否有所提升。圖二十一(A),雙質體轉形策略之二 次轉形株egfp 拷貝數較一次轉形株母體提升了 1.5 至 2 倍,而在單質體轉形策略 之二次轉形株,egfp 拷貝數平均而言較一次轉形株母體提升了 1.5 倍左右,如圖二 十一(B)與表九。
第四章 討論
一 、轉形株表現量差異
轉形株彼此間異源蛋白質表現量有著很大的差異,其中的因素可能有:外源 基因拷貝數目、外源基因插入位置、表觀遺傳(Epigenetics)等等。
1. 外源基因拷貝數目
進行轉形時,進入宿主細胞內的外源基因數目直接影響基因表現量以及異源 蛋白質表現量,因此提高送入宿主細胞內的外源基因數目,對於提高異源蛋白質 產量將有所幫助,不過也有研究指出送入過多的拷貝數反而會造成異源蛋白質產 量降低[74],可能是因為送入過多外源基因導致細胞本身生理功能受到影響。本實 驗室前人利用電穿孔轉形法對菇類進行轉形,可送入多拷貝數的外源基因進入宿 主,使得異源蛋白質表現量達毫克等級[48]。
2. 外源基因插入位置
農桿菌會隨機地選擇位置後將 T-DNA 嵌入宿主染色體,因此利用農桿菌媒介 轉形法進行異源基因表現,外源基因於宿主染色體的插入位置亦是影響表現量的 重要因素,若外源基因插入點位於染色質纏繞緊密的異染色質(heterochromatin)區 域的話,外源基因便不易進行表現,若是插入點位於染色質纏繞較為鬆散的常染 色質(euchromatin)中或是常態表現的基因附近,通常可以得到較高的基因表現量 [75, 76]。
3. 表觀遺傳
此外,表觀遺傳對基因表現亦有著很大的影響,表觀遺傳代表並非由基因序 列突變所引起的基因表現調控現象,例如:組蛋白修飾(histone modifications)或 DNA甲基化(DNA methylation)。過去研究指出甲基化修飾與外源基因表現程度有 所關聯,其中基因啟動子發生CpG甲基化修飾(CpG island methylation),將會抑制 下游基因的表現量,在菇類轉形研究亦指出此現象[76, 77]。有些轉形株即使具有
較多的外源基因拷貝數,但其異源蛋白質含量卻不見得較高,其原因便可能是外 源基因序列受到了表觀遺傳修飾而導致基因表現受到抑制。
二 、轉形株純度
本研究所使用的轉形材料為菌絲塊,該轉形材料的好處是操作方便以及容易 培養,但其面臨的問題則是無法取得純種培養(pure culture)。菌絲塊是一群絲狀真 菌附著於固態培養基上,因此農桿菌媒介轉形法是對著一群絲狀真菌進行轉形,
所產生的轉形株或許有著多種基因型甚至是參雜著野生型菌絲,絲狀真菌無法分 離成單一細胞,因此操作上僅能挑取局部的菌絲並且多次繼代於選擇性培養基以 減少野生型菌絲的比例,如此才能確保轉形株的純度。由於本研究的菌絲體於液 態培養過程並無添加抗生素,野生型菌絲所佔的比例因而提高,為了避免純度問 題影響了結果,各轉形株液態培養於同一天開始,並於同一天收取菌絲,藉此避 免各樣本間純度差異太大。此外如果時間足夠,液態培養過程添加抗生素也是可 行的一個方法。
三 、農桿菌媒介轉形法探討 1. 農桿菌品系
本實驗室先前菇類轉形研究多使用農桿菌LBA4404 以及 AGL-1 進行農桿菌媒 介轉形法,在轉形效率方面兩者並無明顯差異。本研究為了確認不同品系的農桿 菌是否會影響異源蛋白質表現量,而選用了LBA4404 與 GV3101 兩種品系。GV3101 常應用於植物研究,該品系被認為有機會送入較多拷貝數的外源基因至宿主染色
體,因而有較高的異源基因表現量。本實驗結果顯示,相較於利用農桿菌GV3101
所產生的轉形株,LBA4404 產生的轉形株其綠色螢光蛋白質含量比較高,而轉形 效率方面兩者並無明顯差異,因此在美白菇轉形系統之下使用農桿菌LBA4404 會 是比較好的選擇。
2. 共培養條件
農桿菌媒介轉形法係將誘導後的農桿菌與菌絲進行共培養,以將農桿菌的外 源基因轉送至菌絲,如果共培養期間農桿菌過多將會阻礙菇類菌絲的生長,在清
洗的過程也不容易把農桿菌從菌絲塊上去除乾淨,因此適合的共培養條件將有助 於提高轉形率。本實驗將農桿菌接種於50 ml LB 中隔夜大量培養後,使用 IM broth 把菌體懸浮並且調整菌液濃度,通常會調整至OD600 = 1,不過菌液濃度不論是 OD600 = 0.5 或是 OD600 = 2 皆有成功轉形的經驗,而不同菌液濃度的轉形成功率則 需進一步的分析。
四 、重複轉形法探討
本研究的重複轉形第一次轉形株皆為p0390-Cbx-Hiegfp 轉形株,若改以 p0390-AH-Aiegfp 轉形株作為母體進行第二次轉形的話,會有操作不易的問題,原 因是p0390-AH-Aiegfp 轉形株的菌絲比較稀疏單薄,共培養過程農桿菌容易在菌絲 塊上形成菌落,進而導致共培養完後農桿菌清洗不掉使得轉形失敗,如果想對 p0390-AH-Aiegfp 轉形株進行重複轉形,勢必要重新調整農桿菌媒介轉形法的條 件。
本實驗轉形策略分為雙質體轉形策略以及單質體轉形策略,前者係使用不同 的表現載體進行重複轉形,後者則使用同一表現載體進行重複轉形。雙質體轉形 策略的用意是為了證明重複轉形的可行性,利用帶有不同篩選標記的表現載體先 後進行農桿菌媒介轉形,若二次轉形株表現出兩種抗生素抗性,即證實重複轉形 的成功;單質體轉形策略的缺點是無法明確區分一次與二次轉形株,但是不需要 額外的篩選標記便可提升異源表現量,有利於繼續進行重複轉形。比較兩種策略 提升異源蛋白質表現量的能力,雙質體轉形策略最高可將轉形株綠色螢光蛋白質 表現量從75.47 ng/g TSP 提高至 418.83 ng/g TSP,而單質體轉形策略則是提升至 319.58 ng/g TSP,由此結果可得知雙質體轉形策略有較好的效果,推測原因可能是 雙重抗生素的篩選壓力導致外源基因必須持續且大量地表現。但是雙質體轉形策 略的轉形株係培養於雙重抗生素的選擇性培養基,也導致轉形株生長緩慢。
本研究使用即時定量聚合酶鏈鎖反應測定外源基因拷貝數,其中二次轉形株 拷貝數提升量多為一次轉形株的1.5 倍,而非 2 倍或 3 倍,這種非整數倍提升的結 果應是和轉形株純度有所關係,因為轉形株中參雜了野生型菌絲,因而在測定時 降低外源基因拷貝數之數值,使得最終結果低於預期之數值。重複轉形雖然可以 提升異源蛋白質表現量,但是從表九卻發現高表現量的轉形株:T1、O1、O5 其外
源基因拷貝數反而偏低,其原因可能是該轉形株之T-DNA 擾亂了生長相關的基因 導致轉形株生長速率變慢,使得野生型菌絲之比例提高,若需驗證此推論可對蛋 白質表現量高但是外源基因拷貝數低的組別進行插入位置的確認。
第五章 結論與未來展望
本研究主要目的是藉由農桿菌媒介重複轉形法提升菇類轉形系統的異源表現 量,由實驗結果可得以下結論:
1. 以雙質體轉形策略進行重複轉形所得到之二次轉形株可於雙重抗生素的選擇性 培養基上生長,顯示二次轉形株可表現兩種抗藥性,也證實重複轉形之成功。
2. 根據酵素免疫分析的結果,雙質體轉形策略之二次轉形株的綠色螢光蛋白質含
量最高可提升5.5 倍,單質體轉形策略之二次轉形株的綠色螢光蛋白質含量最高
亦可提升4.25 倍。
3. 根據即時定量聚合酶鏈鎖反應的結果,二次轉形株之外源基因拷貝數大多有所 提升,所有的雙質體轉形策略之二次轉形株平均外源基因拷貝數是一次轉形株
的1.66 倍,而單質體轉形策略之二次轉形株平均外源基因拷貝數則是一次轉形
株的1.52 倍,顯示重複轉形確實可以再次把外源基因送入宿主染色體。
以上的結論顯示農桿菌媒介重複轉形法確實可應用於菇類轉形系統,且重複轉形
應可結合其他分生策略,例如刪減後的gpd 啟動子,再更進一步地增強異源基因
表現。
未來,可繼續探討農桿菌媒介重複轉形法,本研究僅進行到第二次轉形,之 後可進一步以單質體轉形策略進行第三次或是更多次的重複轉形並觀察異源蛋白 質是否能更加提升。此外我們還可將農桿菌媒介重複轉形法應用於其他菇類,例 如發展較為完善的金針菇系統,以測試重複轉形在其他菇類可否順利運作。亦可 把重複轉形跟各種分生策略作結合並且表現功能性基因,像是病毒表面抗原或是 醫藥用蛋白質等。
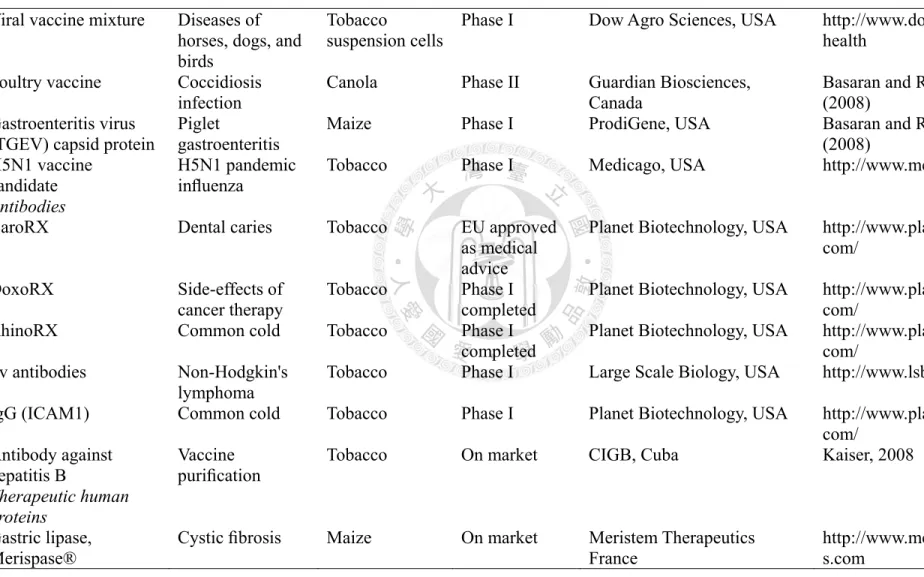
![Table 1. Comparison of heterologous expression system [78]](https://thumb-ap.123doks.com/thumbv2/9libinfo/9600197.628778/42.1263.119.1190.292.714/table-comparison-of-heterologous-expression-system.webp)
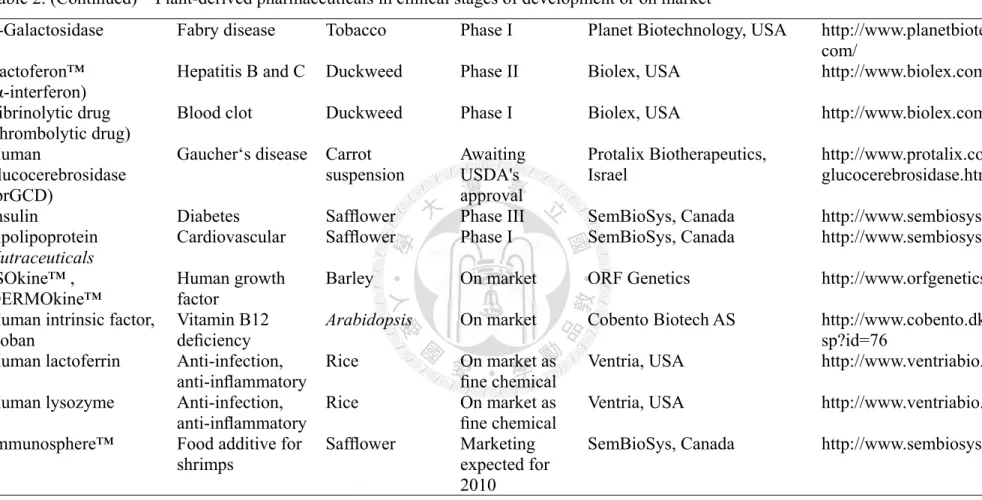
![Table 2. Plant-derived pharmaceuticals in clinical stages of development or on market [79]](https://thumb-ap.123doks.com/thumbv2/9libinfo/9600197.628778/43.1263.106.1188.186.701/table-plant-derived-pharmaceuticals-clinical-stages-development-market.webp)