國 立 交 通 大 學 分子醫學與生物工程所
碩士論文
Functional characterization of the serine threonine kinase
KpnK in Klebsiella pneumoniae CG43
克雷白氏肺炎桿菌 CG43 絲胺酸/蘇胺酸激酶 KpnK
的功能性探討
Student: Hui-Yu Cho
學生: 卓蕙瑜
Advisor: Hwei-Ling Peng, PhD.
指導教授: 彭慧玲 博士 July, 2014
i
論文摘要
近期研究指出,克雷白氏肺炎桿菌之絲胺酸/蘇胺酸激酶 KpnK 會正向調控 細菌抗氧化壓力的反應。本論文探討 KpnK 在肝膿瘍分離株 CG43 中是否還扮演 其他調控角色,首先,我們發現剔除 kpnK 基因後會增加 CG43 對抗巴拉刈的敏 感性,而此突變株並沒有如預期的影響其多醣莢膜的生成,且磷酸甘露醣異構酶 第 98 絲胺酸殘基的磷酸化也不受 kpnK 基因缺損的影響。接著,啟動子活性分 析顯示 kpnK 的表現會受調控胞外結構壓力的雙分子磷酸化系統 CpxAR 正向影 響。有趣的是,kpnK 的基因缺失會增加此菌株凝集酵母菌的活性,此暗示第一 型線毛的表現量會增加;相對的,此基因剔除對 CG43 的胞外組成分子包括第三 型線毛單位蛋白 MrkA 或纖維素的生成量卻無顯著影響。另外,為了證明 KpnK 的酵素活性,還建構了分別帶有 KpnK 定點突變序列(KpnK-S36A, -D201A, -D217A)的三個 pETQ31 重組質體,並以 Pro-Q Diamond 染色分析帶有其中一個 重組質體的 kpnK 基因缺損菌株的總蛋白及重組 KpnK 蛋白的磷酸化狀態,卻發 現二者的磷酸化程度沒有因 kpnK 基因缺損而改變,此結果暗示單一的定點突變 對 KpnK 的活性影響不大。更進一步以 Pro-Q Diamond 染色失去自我磷酸化能力 的截短 KpnK(N-KpnK 或 C-KpnK)和完整 KpnK 的重組蛋白,結果都呈現被磷酸 化的狀態,這些結果顯示其他激酶可能可以代償 KpnK 的磷酸化能力。ii
Abstract
Klebsiella pneumoniae STK, namely KpnK, has recently been shown to positively regulate the oxidative stress response. The thesis investigates if KpnK plays more regulatory roles in K. pneumoniae CG43, a liver abscess isolate. Firstly, the deletion of kpnK gene was found to increase K. pneumoniae CG43 sensitivity to paraquat, however, the mutant did not exert the changes, as suggested by the previous study, on the CPS production or on the phosphorylation state of ManB-S98, the 98th residue serine of phosphomannosemutase. The subsequent promoter activity analysis demonstrated that kpnK was positively affected by the envelope stress regulation two component system CpxAR. It is interesting to note that the kpnK deletion mutant exhibited an increased mannose-sensitive yeast agglutination activity suggesting an increased expression of type 1 fimbriae, by contrast, no apparent changes of the production of the envelope component type 3 fimbriae pillin MrkA or cellulose. Furthermore, the 3 pETQ31-derived recombinant plasmids with sequences coding for the site-directed mutated KpnK (respectively KpnK-S36A, -D201A, and -D217A) were constructed. The analysis via Pro-Q Diamond staining assay revealed that the total cellular proteins or the recombinant KpnK protein of ∆kpnK mutant carrying either of the recombinant plasmids had no change of the phosphorylation pattern, suggesting that the single directed-mutation of the KpnK may have little effect on the KpnK enzymatic activity. Furthermore, the recombinant truncated KpnK proteins, N-KpnK and C-KpnK, which had lost the autophosphorylation activity, remained in phosphorylated form as the recombinant KpnK protein after staining by Pro-Q Diamond. These results suggest that other kinases may compensate the phosphorylation activity of KpnK.
iii
謝誌(Acknowledgement)
碩班時光,說長不長,說短不短。謝謝兩年來許多幫助過我的人,很開心有 這個緣分可以加入溫暖的彭大家庭,讓我在這裡可以無憂無慮的學習。首先感謝 我的指導教授─彭慧玲老師,您宛如媽媽般,指導著我實驗上邏輯的思考,每每 在實驗遇瓶頸的時候,就像燈塔般引領我找到新的方向,也在一次次科學新聞討 論中,教導我們該注意的許多小細節,在這些討論及指導論文修改的過程中,我 總覺得老師的中文、英文以及科學邏輯都好強唷!!(希望我也可以如此),邏輯思 考對我來說是一件要想很久的事,經由老師、學長姐、同學們耐心的訓練,我很 開心我進步了,這一切都要歸功於大家細心的教導。除此之外,老師也時常關心 我們,尤其在我腳受傷的時候,更貼心的給了我護膝,並也給我們充分的機會, 讓我們不僅在實驗上有所成長,在心靈上也有踏實的歸屬感。 也特別感謝中興大學鄧文玲老師以及交通大學梁美智老師,在百忙中撥冗擔 任我的口試委員,用心地給予我修正論文的建議及指點我實驗中遇到的盲點。以 及感謝清華大學張晃猷老師,在我碩二寒假的 joint meeting 及預口試時給予寶貴 的建議,使我有更多方向思考的機會。 感謝實驗室中的夥伴們,因為有你們,我在這裡的生活才能如此絢麗。謝謝 氣質又能幹的靜柔學姊及對實驗超有熱忱的哲充學長,在實驗中我有任何的問題, 你們總是鉅細靡遺的教導我,在實驗卡關的時候,你們也總是絞盡腦汁協助我如 何 trouble shooting,實驗室有你們兩個學長姐的帶領,讓我們可以很快進入狀況。 有趣的冠男學長、美麗的燕曦學姊、以及電腦能力超強的子祥學長,謝謝你們除 了在實驗上的幫助外,並分別在實驗室報帳的事宜、身體健康及如何保養的觀念、 以及幫我處理許多電腦方面的問題上,給予眾多的幫忙,讓我在生活上有更多的 體驗。謝謝我的同伴們─邏輯力超強的偉豐,在我百思不得其解的 pathway 前, 一步一步指導我理出思緒;英文超好的俐君,在我閱讀文獻有問題時給予適當的 協助;個性率直的珍儀,陪我從大學走到研究所 6 年的生活裡,帶領我邁向吃素iv 的境界(雖然我還是會想吃肉),有你們這些可愛的同伴們,除了實驗外,我們一 起去 KTV 瘋、那兩年我們一起去逛的大潤發、中秋節一起烤肉、一起為畢業生 跳的”撐腰”以及許多林林總總的生活大小事,將在我的生命中留下美麗的印記。 學長兼學弟的家睿,謝謝你總在我跟你借摩托車的時候,義不容辭借我;兩位可 愛又天兵的學妹子芸、瑋芝,謝謝你們總在我需要擺 pose 照相的時候,教我裝 一下氣質;以及兩位帥帥的新碩一啟翔、若迪和我一起開心討論著電視劇…等, 實驗室有你們 5 個學弟妹,讓我可將實驗結果的煩惱拋之九霄雲外(雖然最後還 是得要回來面對)。也謝謝張晃猷老師實驗室的 Anita、Clare、芊瑜學姊在緊急 的時刻借我實驗用品以及協助報帳事宜,同學慈芳、韻婷一起修課、討論實驗, 互相切磋成長。有你們的陪伴,讓我在研究之路,獲益良多。 感謝我的大學同學:提示我論文目錄編排技巧的雅妃、在我煩心時提供我遊 玩住宿的楊蓉、怡君、念慈,好感動有你們的陪伴。感謝暨南大學專題實驗室若 梅學姐、椒楹學妹在我舊傷復發的期間,還特別寄那個很厲害的磁石來幫我鎮住 腳傷。以及在大學、研究所間,所有關心指導我的任課老師、學長姐們,真的非 常謝謝你們,讓我在知識的殿堂裡,有更深的探索。 最重要的是感謝對我無怨無悔、不求回報的家人們,每當在我慌張的時候, 總是在我身旁給我力量;在我忙得焦頭爛額時,總是讓我心無旁騖能好好把自己 的目標達成。這輩子能和你們成為家人,是多麼的幸運!!在這重要的時刻,我想 對你們說,我愛你們,雖然孩子不管長多大,父母總有操不完的心,但現在我已 漸漸成熟,請你們放心。最後,我也想謝謝自己,勇敢的以化學系背景接觸生科 領域,並在過程中不斷堅持並汲取新知。雖然即將畢業,在這高度競爭的社會中, 會有更多的挑戰,但我想告訴自己,勇於創新、不害怕嘗試新事物、別等到失敗 才後悔自己還有潛力沒發揮,成功,永遠是留給準備好的人。 中華民國 103 年 7 月 22 日 卓蕙瑜 謹誌
v
目錄
論文摘要... i Abstract ... ii 謝誌(Acknowledgement) ... iii 目錄... v 表目錄... viii 圖目錄... ix 縮寫表... xi 第一章 前言... 1 1.1 克雷白氏肺炎桿菌(Klebsiella pneumoniae, KP) ... 1 1.2 莢膜多醣體(CPS) ... 2 1.2.1 磷酸甘露糖異構酶(ManB)... 2 1.3 蛋白質的磷酸化修飾(protein phosphorylation) ... 3 1.3.1 蛋白 Ser/ Thr 的磷酸化激酶(STK) ... 41.3.2 ESTK─ YihE (RdoA) ... 4
1.3.3 KpnK 的調控及功能 ... 5 1.4 CpxAR 的調控及功能 ... 6 1.5 線毛黏附因子... 6 1.6 抗壓反應(Anti-stress response) ... 8 1.6.1 抗酸機制... 8 1.6.2 抗氧化機制... 9 1.7 研究目標... 10 第二章 實驗材料與方法... 11 2.1 實驗菌株、質體、核酸引子、培養條件... 11 2.2 質體建構... 11
2.2.1 缺失突變體(specific gene-deletion mutants)及回補質體 (complementary plasmid)的建構 ... 11
vi 2.2.2 定點突變體(Site-directed mutagenesis) ... 12 2.3 生長曲線測量... 12 2.4 莢膜多醣體的低速離心定性分析... 12 2.5 巴拉刈(paraquat)及過氧化氫(H2O2)的氧化壓力能力評估 ... 12 2.5.1 生存率測試... 12
2.5.2 紙錠分析(Disc diffusion assay) ... 13
2.6 酸性環境壓力生存率測定... 13
2.7 西方墨點法(Western Blot analysis) ... 13
2.8 蛋白質高量表現之質體建構... 14 2.9 蛋白質大量誘導... 14 2.10 Pro-Q diamond 染色分析蛋白質的磷酸化 ... 15 2.11 酵母菌凝集測試(Yeast Agglutination) ... 15 2.12 啟動子(Promoter)分析 ... 15 2.12.1 啟動子報告載體建構... 15 2.12.2 啟動子活性分析... 16 2.13 生物膜染色分析... 16 2.14 剛果紅(Congo red)及鈣螢光(Calcofluor)表現型分析 ... 17 2.15 數據分析... 17 第三章 實驗結果... 18 3.1 K. pneumoniae CG43S3 ∆kpnK_1 的 CPS 生合成... 18 3.2 kpnK 下游基因 dsbA 缺失對 CPS 生合成的影響 ... 18 3.3 CG43S3 ∆kpnK 基因缺損株的建構與其生長曲線分析 ... 18 3.4 kpnK 基因缺損突變株中 ManB 的磷酸化 ... 19 3.5 kpnK 基因缺損株的抗氧化能力分析 ... 19 3.6 以 LacZ 報導系統探討 CpxAR 對 kpnK 基因的調控 ... 19 3.7 kpnK 基因缺損株的抗酸能力 ... 20 3.8 kpnK 缺損突變株凝集酵母菌的活性分析 ... 20
vii 3.9 kpnK 基因缺失增加第一型線毛單位蛋白 FimA 的生成 ... 21 3.10 kpnK 基因缺失對第三型線毛表現及生物膜形成能力的影響 ... 21 3.11 kpnK 基因缺損株的 EPS 表現 ... 22 3.12 KpnK 定點突變質體的選殖與建構 ... 22 3.13 KpnK 定點突變對其酵素活性的影響 ... 22 3.14 在 CG43S3 中測試 N-及 C-KpnK 的磷酸化 ... 23 3.15 N-KpnK 及 C-KpnK 在 kpnK 缺失突變株中激酶活性的探討 ... 23 3.16 在 KPCG43 基因體中有多套 STK 基因 ... 23 第四章 討論... 25 第五章 參考文獻... 29
viii
表目錄
表一、本研究所使用與建構的菌株... 43 表二、本研究所使用與建構之質體... 45 表三、本研究所使用之核苷酸引子... 47
ix
圖目錄
圖 一、kpnK_1 基因缺損突變株在 K. pneumoniae CG43 中的 CPS 生合成 ... 49 圖 二、基因缺損突變株之建構示意圖及低速離心觀察∆kpnK_1、∆dsbA 及 ∆kpnK_1∆dsbA 在 K. pneumoniae CG43 中的 CPS 生合成 ... 51 圖 三、在 K. pneumoniae CG43 中建構 kpnK 基因缺失突變株及回補株 ... 52 圖 四、K. pneumoniae CG43 中的 kpnK 基因缺損突變株及回補株在 LB 及 M9 培 養液中的生長曲線分析... 53 圖 五、在 K. pneumoniae CG43 中 kpnK 基因缺損突變株的 CPS 生合成及 ManB 的磷酸化... 54 圖 六、在 K. pneumoniae CG43 中 kpnK 基因缺損突變株對抗 H2O2及 paraquat 抗 氧化能力分析... 55 圖 七、在 K. pneumoniae CG43 中 CpxR 對 kpnK 基因的轉錄調控 ... 56 圖 八、kpnK 基因缺損突變株在 K. pneumoniae CG43 中的抗酸能力 ... 57 圖 九、kpnK 基因缺損突變株在 K. pneumoniae CG43 中凝集酵母菌的活性分析 ... 58 圖 十、在 K. pneumoniae CG43 中 kpnK 基因缺失對第一型線毛表現的影響及轉 錄調控... 60 圖 十一、在 K. pneumoniae CG43 中 kpnK 基因缺失對第三型線毛表現的影響 61 圖 十二、在 K. pneumoniae CG43 中 kpnK 基因的缺失對生物膜形成能力的影響 ... 62 圖 十三、在 K. pneumoniae CG43 中 kpnK 缺損突變株的纖維素表現 ... 63 圖 十四、kpnK 在 K. pneumoniae CG43 及 E. coli 中胺基酸序列比對分析 ... 64 圖 十五、在 K. pneumoniae CG43 中 S36A、D201A、D217A 的定點突變質體建 構及定序確認圖... 65 圖 十六、以 Pro-Q Diamond 磷酸化蛋白質螢光染劑分析 K. pneumoniae CG43 中x
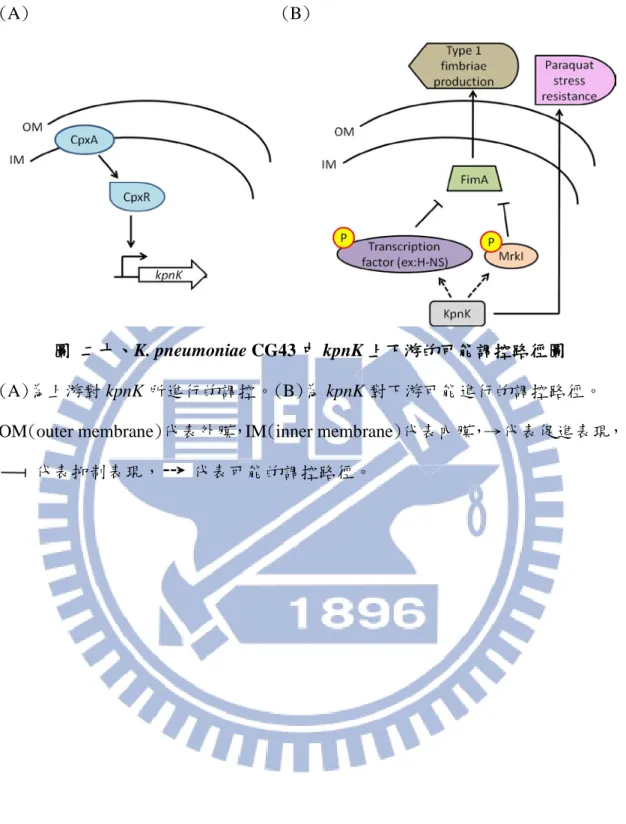
KpnK-S36A, -D201A, -D217A 定點突變對其酵素活性之影響 ... 66 圖 十七、在 K. pneumoniae CG43 中分析 N-KpnK 及 C-KpnK 磷酸化之變化 ... 68 圖 十八、在 K. pneumoniae CG43 ∆kpnK 菌株中分析 N-KpnK 及 C-KpnK 磷酸化 之變化... 69 圖 十九、K. pneumoniae CG43 中共有 4 套 STK 基因 ... 70 圖 二十、K. pneumoniae CG43 中 kpnK 上下游的可能調控路徑圖 ... 71
附錄 一、在 K. pneumoniae CG43 中 manB 的缺失突變體的 CPS 生合成及 ManB S98 影響其酵素活性 ... 72 附錄 二、在 KP NTUH-K2044 之磷酸蛋白質體(phosphoproteome)分析... 73
xi
縮寫表
ATP adenosine triphosphateBCIP 5-bromo-4-chloro-3-indolylphosphate bp base pair
CFU colony forming unit(s)
CIAP calf intestine alkaline phosphatase CPS capsular polysaccharide
DNA deoxyribonucleic acid DTT dithiothreitol
EDTA ethylenediamine-tetraacetic acid ESBL extended-spectrum β-lactamase
IPTG isopropyl-1-thio-β-D-galactopyranoside kb kilobase(s)
kDa kilodalton(s)
KpnK Klebsiella pneumoniae Kinase LB Luria-Bertani μg microgram mM milligram μM micromolar mM Millimolar ManB phosphomannomutase NBT nitro blue tetrazolium chloride
ONPG ortho-nitrophenyl-β-D-galactopyranoside PAGE polyacrylamide gel electrophoresis PCR polymerase chain reaction
PVDF polyvinylidene difluoride SDS sodium dodecyl sulfate
ST(P)K serine/ threonine (protein) kinase TCS two-component system
1
1.1 第一章 前言
1.1 克雷白氏肺炎桿菌(Klebsiella pneumoniae, KP) KP 是一株伺機性感染的革蘭氏陰性菌,屬腸內菌科,具有厚重莢膜、不具 鞭毛、為一不具移動性的兼性厭氧菌。除了自然環境外,KP 在健康人體如:口 腔黏膜、眼睛、腸道、尿道,以及肺部等黏膜部位也常被發現(Ryan et al., 2004)。 在免疫不全的病人身上,常導致菌血症、敗血症、眼內炎、腦膜炎、肺炎、尿道 以及呼吸道感染;而在亞洲,KP 已成為引發肝膿瘍的首要原因,在 2004 年,更 有超過 900 個案例在台灣被報導,如不加以治療,則有高達 90%的致死率(Fang et al., 2004)。另外,KP 也是造成群聚感染以及院內感染常見的病原菌之一 (Podschun et al., 1998)。 近年來,由於抗生素的濫用,導致抗藥菌株不斷的出現。1989 年,第一株 帶有多重抗藥性 extended-spectrum-beta-lactamase(ESBL)基因的 KP 在美國被發 現後,多重抗藥的 KP 菌株隨即在美國境內快速傳播(Colodner et al., 2004); Carbapenems 原為用來管控多重抗藥性菌株感染之藥物,但在 2008 年,抗 Carbapenems 的 KP 菌株 KPC 出現(French et al., 1996; Hirsch et al., 2010;Nordmann et al., 2009),接著,更有 NDM-1 KP 的出現,至今,所有帶有 NDM-1 基因的分離株皆有多重抗藥性(Kumarasamy et al., 2010; Yong et al., 2009),因此, 如何管控 KP 所引發的感染,遂成了一項嚴重的挑戰。
KP 的致病因子包含包含酯多醣(lipopolysaccharide, LPS)、攝鐵系統(iron acquisition system)、分泌系統(secretion system) 、黏附因子(adhesion)、莢膜多 醣體(Capsular polysaccharides, CPS)。而本實驗室所研究的 KPCG43 為長庚紀念 醫院從糖尿病患者的肝膿癢分離株,因其表面受厚重的 CPS 包覆,菌落圓潤光 滑並具有高度黏滯性(Chang et al., 1996);在 KPCG43 中有 8 套攝鐵系統(Lin et al., 2011),可幫助細菌得以在缺鐵的環境或宿主中增生;另外,第三型線毛為外 膜上的髮狀構造,是其主要黏附因子,有助其黏附於生物或非生物器材的表面形
2 成生物膜 (Huang et al., 2006)。 1.2 莢膜多醣體(CPS) 莢膜多醣體可使細菌躲避補體系統(Complement system)攻擊且具有抗吞噬 (anti-phagocytosis)的能力,目前被分離出的莢膜血清型約 77 種,其中以 K1 及 K2 毒性最高,也是造成肝膿瘍主要的血清型(Wacharotayankun et al., 1992)。其莢 膜多醣體構成單元為 [→4)-Glu-(1→3)-α-Glc-(1→4) -β-Man-(3→
1)-α-GlcA)-(1→]n。負責 K2 莢膜生合成的 gene cluster 包含 19 個開放讀框(open reading frames),分屬 3 個轉錄單位,其中有 4 個基因 wzi-wza-wzb-wzc 與 E. coli 的相對基因具有高度序列相似性。Wzi 是一外膜蛋白,在∆wzi 中,可以明顯看 到多糖莢膜大量減少,暗示著 Wzi 參與連接多醣莢膜於細胞表面的工作;嵌在 外膜上的 Wza 是一通道蛋白,與嵌在內膜上的酪胺酸激酶(protein tyrosine kinase, PTK)Wzc 交互作用,而將膜間的多醣莢膜送出外膜;Wzc 酪胺酸激酶能自我磷 酸化或磷酸化其他蛋白,而 Wzb 酪胺酸去磷酸酶(protein tyrosine phosphatase, PTP)可去除酪胺酸上共價接合的磷酸根,而 PTKs 及 PTPs 的基因多半坐落與製 造及運輸 CPS 有關的基因座上(Cozzone et al., 2009)。我們實驗室也發現 KP CG43cps 基因組的表現,可受 Wzc 磷酸化所調控,其中,磷酸甘露糖異構酶 ManB 可能是 Wzc 磷酸化的標的(萬舉豪,2012)。
1.2.1 磷酸甘露糖異構酶(ManB)
ManB 具有雙重活性的酵素可以催化 CPS 的前驅物 mannose-1- phosphate 與 mannose-6-phosphate 之間轉換的可逆反應;此外,ManB 也可催化
glucose-1-phosphate 與 glucose-6-phosphate 間之轉換,因此此酵素被稱為
phosphomannomutase 或 phosphoglucomutase (Goldberg et al., 1993)。manB 基因 座落於多醣莢膜生合成基因組上,與相鄰的基因產物共同參與多醣莢膜的生合成。 Pseudomonas aeruginosa 的 ManB 酵素的絲胺酸(Ser108)在催化
3
應中先將第 108 絲胺酸磷酸化,然後以共價鍵接在 glucose-1-phosphate 的 6 號碳 上,形成 glucose-1,6-bisphosphate 中間產物,再經由分子翻轉使 1 號碳接近 Ser108 並以共價鍵結磷酸根,最後釋出產物 glucose-6-phosphate (Regni et al., 2006)。 我們實驗室的萬舉豪藉由分析序列結果,推測 KPCG43 ManB 的 Ser 98 之磷酸化 可能影響其酵素活性,而其磷酸化被一未知的激酶所調控(萬舉豪,2012)。 1.3 蛋白質的磷酸化修飾(protein phosphorylation) 細菌的蛋白質形成後修飾主要包含:乙醯化(acetylation)、甲基化 (methylation)、蛋白酶解(proteolytic degradation)、脂化(lipidation)以及磷酸化 (phosphorylation)。其中,磷酸化通常發生在絲胺酸(Ser)、蘇胺酸(Thr)、酪胺 酸(Tyr)、以及組胺酸(His)的殘基上。其磷酸化系統可分為雙分子調控系統 (Two-component system,TCS)、磷酸轉移系統(phosphotransferase system, PTS) 和三磷酸腺苷依賴系統(ATP-dependent kinase phosphorylation system)。TCS 包 含兩個功能獨特的蛋白質 sensor kinase 受到外界訊息刺激後,藉由消耗 ATP, 將磷酸激團以共價鍵接在自身的組胺酸殘基上,經自我磷酸化後,再將磷酸基 傳給另一蛋白 response regulator 的天門冬胺酸殘基,而此 response regulator 的 磷酸化與否決定其調控功能。PTS 同樣將磷酸基團接在組胺酸上,與 TCS 最大 的不同在於 PTS 的磷酸基團由 phosphoenol-pyruvate (PEP)所提供,而且其磷酸 基團經一連串的蛋白質傳遞後,最終傳到醣分子(van Tilbeurgh et al., 2001)。在 第三個系統中,磷酸化位置發生於蛋白質的 Ser、Thr 以及 Tyr。絲胺酸/蘇胺酸 激酶(STK)和 TCS 的不同在於:(一)形成不同的磷酸酯類:STK 形成
phosphoester 而 TCS 形成 phosphoramidate (Stock et al.,2000);(二)不同數目的 磷酸根傳遞:TCS 只傳遞一個磷酸根,STK 卻可能因多種激酶參與磷酸化,而 有多重磷酸根傳遞的現象(Inouye et al., 2008; Stock et al., 2000);(三)TCS 影響 基因的轉錄(transcription),而 STK 除了轉譯後修飾影響蛋白的活性外,當其磷 酸化到標的蛋白為轉錄因子時,也會因而影響其下游基因的轉錄(Lin, 2010)。
4
1.3.1 蛋白 Ser/ Thr 的磷酸化激酶(STK)
真核細胞的蛋白磷酸化系統已被廣泛研究(Bakal et al., 2000),而至最近幾十 年間,才陸續有細菌 STK 的報導,而某些和真核 STK 序列具有高度相似性的一 群被稱為類真核型的絲胺酸/蘇胺酸激酶(Eukaryotic-like Ser/ Thr Kinase,
ESTK)(Hanks et al., 1995),但並非所有細菌 STK 都為 ESTK。
典型的 ESTK 和真核細胞中的 STK 催化中心具有高度的結構相似性(Canova et al., 2014),且符合“Hanks’ motifs”法則:通常 N 端由較小區域的β-sheets 組 成,主要和磷酸根的供應者 ATP 結合,為可能的 ATP 鍵結區(ATP binding site); C 端由較大範圍的α-helixes 組成,會辨認並結合受質的特定序列殘基,為受質 鍵結區 (Pereira et al., 2011)。ESTK 具有 Glycine-rich motif 和 ATP 結合(但有些 ESTK 並無此特徵),當 Glycine-rich motif 缺失時,此激酶可藉由 lysine-rich motif 或 Glutamate-rich motif 和 ATP 鍵結以獲得高能磷酸根;另外,ESTK 具有和金屬 (Mg2+、Mn2+)鍵結的 DFG motif,以穩定激酶活性;在 DFG 及 PE motif 間為磷 酸化修飾的活化部位,PE 的 Glutamate 會和 Arginine 形成鍵結;而 RDXXXXN motif 為鍵結 Aspartate,形成 ATP 傳遞予受質結合的結構。
首先在 E. coli 中發現參與檸檬酸循環酵素 IDH (isocitrate dehydrogenase)的 活性與其 Ser/ Thr 磷酸化有關(Garnak et al., 1979),然而,IDH 是透過雙功能的 激酶/去磷酸酶(bifunctional kinase/phosphatase) 在 Ser 的位置進行磷酸化,此雙 功能的激酶/去磷酸酶和 ESTK 的胺基酸序列並不相似(La Porte et al., 1985)。直 到 1991 年,第一個名為 Pkn1 的 ESTK 在 Myxococcus xanthus 中被發現,其激脢 活性會影響生長(Munoz-Dorado et al., 1991)。在其他的細菌中,陸續也發現 ESTK 與其代謝,胞子、細胞壁及莢膜的生成有關,也可能是細菌的毒性因子
(Edwards-Jones et al., 2004; Richards et al., 2006; Roque et al., 2009; Zheng et al., 2007)。
1.3.2 ESTK─ YihE(RdoA)
5
似性。yihE和其下游基因dsbA為一組基因操縱子(operon),此操縱子受Cpx的轉 錄調控(Turcot et al., 2001; Suntharalingam et al., 2003)。而yihE (或rdoA)相關的表 現型,可能會因為在不同種細菌間有所不同(Suntharalingam et al., 2003; Lin, 2010)。許多相關報導yihE/rdoA的缺失突變會影響基因的轉錄、表現量、以及磷 酸化其他蛋白的能力:S. flexneri的微陣列(microarray)分析實驗發現rdoA剔除後 會影響和能量代謝、碳原子降解、胺基酸及核苷酸的合成及蛋白質運輸相關之基 因的轉錄(Li, et al., 2001);也有研究提出DnaK、GroEL、Ef-Tu具有RdoA依賴性 磷酸化的現象,因此推測它們是RdoA的受質(Roque, 2009),但仍未經證實。S. flexneri YihE(RdoA)會調控galERKM表現而影響LPS的生成量,進而決定其毒 性高低(Edwards-Jones et al., 2004);但在S. typhimurium中,YihE(RdoA)並不 影響LPS的表現量;另外,RdoA也會影響線毛的生成或造成鞭毛狀態的改變 (Richards, 2006; Zheng et al., 2007)。
結構分析的研究結果顯示 YihE(RdoA)在胺基酸 H199 的位置,會藉由和 其他胺基酸的交互作用改變結構,進而影響激脢活性;H199 和 D201 及 V216 的 主鏈形成氫鍵鍵結,使得 D201 和 D217 的支鏈方向改變,而 D217 的主要功用 是和鎂離子鍵結,進而催化其受質的羫基(hydroxyl group)去質子化 (deprotonation),而受質取得 ATP 的γ-磷酸根後會被磷酸化,因此,將 D217 突 變成 alanine 則會喪失其激酶的活性;另外,S36 會透過其羫甲基(hydroxymethyl group)和 ATP 的β-磷酸根結合,形成過度態來穩定 ATP(Zheng et al., 2007)。
1.3.3 KpnK 的調控及功能
2012 年,類真核型磷酸化激酶 KpnK (K. pneumonia Kinase)首先在 KP MGH78578 中被證實此激酶會受外界環境壓力的刺激,影響 KP 抗氧化及抗抗生 素的生存率(Srinivasan et al., 2012)。KpnK 不具完整的特定 ESTK 胺基酸序列, 但仍被歸屬於 APH-3’-IIIa and choline kinase 家族,具有鍵結 ATP、錳離子相關 序列,富含賴胺酸(lysine-rich)和 RDX6N motis,屬於非典型的 ESTK。
6
在kpnK的基因缺失會使KP對抵抗滲透壓力、氧化壓力及抗生素的能力降低;同 時,KpnK在轉錄過程中會受CpxR的正向調控(Srinivasan et al., 2012)。
1.4 CpxAR 的調控及功能
Cpx是一雙分子訊息傳遞系統(two component system,TCS),由CpxA (sensor kinase)及CpxR (response regulator)組成。會受胞外結構壓力(Envelope stress)的 刺激而提高表現,Envelope stress包含胞外酸鹼度、氧化還原壓力、化學分子濃 度造成滲透壓變化、外膜酯蛋白或膜間質(periplasm)中摺疊錯誤蛋白的堆積等 (Keller and Hunke, 2009; Nakayama and Watanabe, 1995; Snyder et al., 1995)。 CpxA受刺激活化後將訊號傳遞給CpxR,CpxR是轉錄因子,參與蛋白質折疊、外 膜結構以及毒性因子等基因的轉錄調控(Humphreys et al., 2004; Nevesinjac and Raivio, 2005; Price et al., 2009; Stefanie et al., 2011)。Cpx的活化可增加CpxAR和 CpxP的表現,CpxP為CpxA的負向調控子,當外界無刺激時,CpxP直接和CpxA 鍵結而抑制其下游途徑的調控;當膜間質中出現了摺疊錯誤的蛋白時,CpxP抑 制CpxA的能力減弱,隨之,與折疊錯誤的蛋白結合後,交給DegP水解酶將折疊 錯誤的蛋白水解(Raivio et al., 1999)。 Cpx 會藉由抑制 csgBADEFG 而影響線毛的表現,也可抑制鞭毛的 motor 蛋 白活性與其他如:motABcheAW、tsr 及 aer 等可影響細菌移動之基因的表現;相 對的,CpxR 也可正向調控基因的表現,譬如:CpxR 可促進合成二級訊息分子 cyclic di-GMP 的 diguanylate cyclase YdeH 的基因表現,進而影響細菌的移動及 生物膜(biofilm)的生合成(Stefanie et al., 2011)。
1.5 線毛黏附因子
KP 外膜的主要結構組成包括 CPS、LPS 和黏附因子第一型和第三型線毛 (type 1 and type 3 fimbriae),第一型和第三型線毛幫助 KP 黏附人體的呼吸道或 尿道得以形成菌落後進一步造成感染。線毛由主要蛋白 major pillin 聚合形成線
7
狀,這些單位蛋白在膜間質中被 chaperone 保護,免於被蛋白水解酶降解,在外 膜上的 usher protein 為基座,可啟動單位蛋白的聚合,而在細菌表面形成線毛。 第一型線毛是大腸桿菌造成尿道感染決定性的黏附因子(Connell et al., 1996),具 和被甘露糖 mannose 糖基化的蛋白專一性的鍵結能力,其組成基因序列、基因結 構以及調控路徑和 KP 的第一型線毛有很高的相同性(Struve et al., 2008; Clegg et al., 1987; Gerlach et al., 1989; Schembri et al., 2005)。大腸桿菌第一型線毛由 fimAICDEFGH 構成,主要的單位結構蛋白為 FimA,次要蛋白由 FimI、FimF 和 FimG 組成,FimH 為位於線毛頂端的黏附蛋白, FimC 保護蛋白及 FimD 基座蛋 白共同參與線毛的聚合作用。KP 的第一型線毛基因組在 fimAICDEFGH 後還有 fimK 基因, FimK 可以活化 KP 第一型線毛的表現(Wang et al., 2013)。FimB 及 FimE 具有 DNA 重組酶的能力,負責調控第一型線毛的表現,KP 第一型線毛 表現量較少,可能是由於厚重莢膜和線毛的交互表現影響(Schembri et al., 2005; Matatov et al., 1999; Sahly et al., 2000),也可能是 KP 在胞外主要表現第三型線毛 所致(Wu et al., 2012)。
第三型線毛可凝集經單寧酸(tannic acid)處理過的紅血球,此外,第三型線 毛受 c-di-GMP 調控且和 Biofilm 的生合成有關(Wu et al., 2012);第三型線毛和 導尿管相關的尿道感染有很密切的關係即可能是形成生物膜所致(Warren et al., 2001;Maki et al., 2001)。第三型線毛 mrkABCDF 操縱子(operon)轉譯的主要結構 蛋白為 MrkA、MrkF 為次要結構蛋白、MrkD 為黏附蛋白,三者均有助於生物膜 的生成(Huang et al., 2009);MrkB 及 MrkC 分別為保護蛋白及基座蛋白;mrkF 下游 mrkH、mrkI 與 mrkJ 構成決定第三型線毛活性的調控基因組,MrkH 為一 PilZ domain 蛋白,可與 c-di-GMP 鍵結;MrkI 為一 LuxR 型的轉錄因子;MrkJ 為一解環化 c-di-GMP 的 phosphodiesterase (吳健誠,2010)。在 KPCG43 中已 發現 mrkI 或 mrkH 的基因缺失會降低 MrkA 的啟動子活性及蛋白生成,而高量 表現 MrkH 時會顯著增加第三型線毛的表現;另外,mrkJ 基因缺失會些微增加 MrkA 的表現(鄭威芸,2011;Wu et al., 2012)。
8 1.6 抗壓反應(Anti-stress response) 細菌須克服各式各樣的壓力,以能生存在這個環境中。常見的壓力來源,包 括:環境中不同的酸鹼度(pH)、滲透壓、以及氧化壓力。細菌在感染人體後,首 當其衝的是胃酸所造成的影響,而通過腸道時,則需要耐受鹼性的消化液,以及 高度的滲透壓力,為了適應這些嚴峻的環境,細菌發展出一些特定的生存機制, 以保護生物分子結構免受破壞。 1.6.1 抗酸機制 在低酸鹼值下,胃幽門桿菌產生大量的尿素分解酵素(urease)消耗氫離子, 將 ammonia 轉換成 amine,以中和細胞質及膜間質的酸鹼度;此外,胃酸刺激也 可影響其泳動性,使胃幽門桿菌移動至胃黏膜上皮細胞的近中性區(Boot et al., 2002)。大腸桿菌以不同的抗酸機制通過胃酸,志賀氏菌(Shigella flexneri)和大腸 桿菌一樣擁有強的抗酸能力,相對的,霍亂弧菌(Vibrio cholerae)的抗酸能力較 低(Merrell et al., 2001)。
大腸桿菌有 5 種抗酸途徑為 AR(acid resistance)1~AR5(Foster et al., 2004; Zhao et al., 2010)。大腸桿菌於 pH5.5 LB 培養至穩定生長期(stationary phase)後 換置於 pH2.5 的限制培養基(minimal media),即可啟動 AR1 抗酸系統,AR1 的 活性受σS 與 CRP(cAMP receptor protein)調控,並受葡萄糖抑制;AR2~AR5 都 是以胺基酸去梭酸基酵素(amino acid decarboxylase)消耗特定胺基酸與氫離子, 產生終產物胺類(amines)與二氧化碳而提高 pH 值;一般而言,厭氧的酸性環境 能夠誘導去梭基酵素的作用,AR2 及 AR3 作用於 pH2.5 的強酸環境;AR4 及 AR5 的抗酸系統則與細菌在 pH4.5 環境下的生存有關(Kashiwagi et al., 1992)。 革蘭氏陰性菌的抗酸系統,除了細胞質中的去羧酸基酵素外,膜間質的伴隨 蛋白也能夠辨識酸刺激而保護蛋白質使其維持活性並避免蛋白沉澱或堆積。大腸 桿菌參與膜間質抗酸反應的伴隨蛋白為 HdeA 及 HdeB,此二基因位於同一 operon, 受酸誘導表現(Kern et al., 2007)。阻斷 HdeAB 表現可明顯降低大腸桿菌、痢疾 桿菌或布氏桿菌(Brucella abortus)在酸壓力下的存活率(Malki et al., 2008;
9
Valderas et al., 2005);恢復 HdeA 或 HdeB 任一功能,可增進大腸桿菌存活率。 HdeA 能結合的膜間質蛋白包含轉運蛋白、代謝酵素、伴隨蛋白、酯蛋白及蛋白 質分解酵素等 (Zhang et al., 2011)。 KP 與大腸桿菌同屬於腸內菌科,但缺乏 AR2 及 AR3,在 pH3 的存活率極 低。在 KPCG43 中,TCS 的調控基因 rcsB 的缺損,可導致其在 pH3 的存活率降 低 30%,進一步以二維電泳分析 RcsB 基因缺損株於酸性環境下的蛋白質體變化, 發現表現量改變幅度最高的蛋白質為 YfdX,而 YfdX 的表現決定其在靜態培養 條件下的抗酸反應(林志桓,2009);而 YfdX 在酸性環境具備伴隨蛋白的活性, 可保護膜間質蛋白免於發生沉澱反應(樊力成,2012);另外,剔除 KP CG43 的 hdeB,可降低其在 pH3 的存活率降低,HdeB 的伴隨蛋白活性也被證實(曾品瑄, 2011);然而,剔除 KPCG43 尿素酵素基因並不影響其於酸性環境下的存活率(林 芳瑜,2013),但可降低 KP 在腸胃道中與野生菌株的競爭力(Maroncle et al., 2006)。 1.6.2 抗氧化機制 在感染的過程中,細菌的抗氧化防禦機制對於其毒性扮演著重要的角色。氧 化壓力主要是由 reactive oxygen species(ROS)和 reactive nitrogen species(RNS)所 造成,ROS 包含超氧自由基(superoxide anion, O2•¯)、過氧化氫 (hydrogen peroxide, H2O2)以及氫氧自由基(hydroxy1 radical, OH˙),而 RNS 則包含了帶 電的一氧化氮(nitric oxide radical, NO˙)及過氧化氮(peroxynitrite, ONOO─
)。 細菌體內 DNA 的受損,以及折疊錯誤的蛋白質、脂質都會對細菌造成氧化壓力, 進而影響細菌的生長及繁殖。而在感染過程中,細菌處在一個充滿氧化壓力的環 境,這時氧化壓力的來源主要來自(一)細菌本身體內的氧化壓力:例如電子傳 遞鏈(electron transport chain, ETC);(二)外在環境:除了環境中原本就存在的 氧化還原反應所造成的氧化壓力外,在感染宿主的過程中也會出現競爭的細菌, 這時競爭者也會產生 ROS 來與之競爭;(三)宿主體內的吞噬細胞 (phagocytic cells):細菌進到宿主體內之後,受到吞噬細胞的辨識,而將細菌吞進細胞裡,
10
吞噬細胞裡的 NADPH oxidase 會與氧氣作用而形成大量的超氧自由基及過氧化 氫,來攻擊入侵的細菌(Imlay, 2003)。
在這些氧化壓力下,細菌酵素 SodA、SodB、SodC、KatE、KatG 可清除氧 化自由基以免受到傷害(Yang et al., 2010; Dong et al., 2010; Cruz-Ramos et al., 2002)。KP CG43 ΔkatG 對 H2O2處理敏感,KP CG43 sodA 及 yjcC 的基因缺失突 變會增加對 paraquat 的敏感性,這些結果顯示 SodA、YjcC 或 KatG 可能是 KP 抗氧化壓力的反應因子;另外,kvgA 的缺失突變體,會降低 sodC 與 katG 的表 現;而 rpoS 的缺失,也會降低 kvgAS 的表現量(吳宛怡,2012;黃靜柔,2013)。 1.7 研究目標 根據前人的研究(2012)顯示,ManB 扮演決定莢膜生合成的重要角色,而 ManB-Ser 98 可被未知的磷酸激酶所磷酸化,此位置的磷酸化可能影響其酵素活 性。本實驗目的則在於證實 KpnK 是否為 ManB-Ser 98 的磷酸激酶及其抗氧化的 活性,同時,確認 KpnK 是否受 CpxAR 所調控及可能影響其表現的因子;進一 步探討其可能影響胞外組成分子表現的調控功能:確認 kpnK 基因剔除是否影響 胞外主要組成分子 CPS、第一型及第三型線毛的生合成、抗酸能力反應及生物膜 形成能力;最後,也將驗證 KpnK 重組蛋白的磷酸激酶活性。
11
第二章 實驗材料與方法
2.1 實驗菌株、質體、核酸引子、培養條件
實驗用菌株、質體、核酸引子分別詳列於表一、表二、表三。用於鏈聚合酶 連鎖反應(PCR)中,作為增幅目標基因之模版之核酸,源自於林口長庚紀念醫院 臨床分離株 K. pneumonia CG43(K2),分離自肝膿瘍病患(Peng et al., 1992),在 小鼠體內具有很高的致命性(Chang HY et al., 1996)。實驗中,細菌皆培養於 37°C、 Luria-Bertani (LB)液態培養基或固態 LB 洋菜膠培養基上,並依不同使用條件加 入不同濃度之抗生素包括 kanamycin(25 μg/ml)、ampicillin(100 μg /ml)、
tetracycline(5 μg/ml)、streptomycin(500 μg/ml)以及 cloramphenicol(35 µg/ml)。 細菌的生長狀態則藉由波長 600 奈米(nm)光吸收度(OD600)所測量。
2.2 質體建構
2.2.1 缺失突變體(specific gene-deletion mutants)及回補質體
(complementary plasmid)的建構
KP CG43 基因體上的缺失突變及回補,是將特定目標基因的前後約
~800-1000 bp 的片段,選殖(clone)入 pKAS46 這個自殺型載體,因此當 KP CG43 在複製時,會將攜帶的片段置換入基因體上,且由於 pKAS46 含 rpsL,空載體 (vector only)會被鏈黴素(streptomycin)所淘選(selection)掉。建構完成的質體會 由 E.coli S17-1λ pir 以 conjugation 的方式送入 KP CG43,之後則會與基因體進行 同源互換(homologous recombination),再以 M9 kanamycin(25 μg/ml)與
ampicillin(100 μg /ml)固態洋菜膠培養基進行篩選,所得菌落再以
streptomycin(500 μg/ml)固態 LB 洋菜膠培養基進行篩選,則會得到同時對 kanamycin 與 ampicillin 型以及抗 streptomycin 型的菌落,即為所求突變體或基因 體回補。最後,再以 PCR 的方式確認。
12
選殖(clone)入 pRK415 的載體中,(不包含啟動子的質體之後實驗時,須加 IPTG 以誘導表現),建構完成的質體會由 E.coli S17-1λ pir 以 conjugation 的方式送入 KP CG43,再以 M9 tetracycline(5 μg/ml)固態洋菜膠培養基進行篩選,所得菌落 再以 streptomycin(500 μg/ml)及 tetracycline(5 μg/ml)固態 LB 洋菜膠培養基進行 篩選,則會得到同時抗 tetracycline 及抗 streptomycin 型的菌落,即為所求質體回 補菌落。其中,pRK415-stk 和 pRK415-kpnK 位分別代表包含及不包含啟動子 (promoter)的 KpnK 質體型回補。 2.2.2 定點突變體(Site-directed mutagenesis)
定點突變以 yT&A-HY5 為板模,並以 Thermo scientific Inc.的 Phusion High-Fidelity DNA Polymerase kit 分別製作了 S36A、D201A、D217A、T325A, 實驗流程皆參考操作手冊(Thermo scientific Inc.)。再以 BamHI、SacI(選購自 MBI Fermentas)酵素切下,並與 pETQ31 相接。實驗所需之核酸引子,皆由 MDBio, Inc. (Taiwan)及 IDT, Inc. (Taiwan)所合成。建構的所有質體詳列於表一及表二。
2.3 生長曲線測量 菌株在 LB 中培養 16 小時(overnight),將 20µl 的菌液以 200 倍稀釋加入 4 ml 的 LB 中,並以 37°C、搖動培養,在特定時間點記錄 OD600的吸光值以得到生長 曲線圖。呈現的數據為三次獨立實驗中具代表性的一次。 2.4 莢膜多醣體的低速離心定性分析 菌株在 LB 中培養 16 小時(overnight),再以 5000 rpm,3 分鐘低速離心, 並將結果以照相呈現。 2.5 巴拉刈(paraquat)及過氧化氫(H2O2)的氧化壓力能力評估 2.5.1 生存率測試 細菌在 LB 培養液 37°C 培養 16 小時後,以 40 倍稀釋至 LB,在培養至 OD600
13 約 0.6~0.8 左右,取 1 ml 菌液至微量離心管,以 15000 rpm 離心 5 分鐘後去掉上 清液,再加入 1 ml 濃度為 3 mM 的巴拉刈(paraquat, PQ)或 10 mM 的 H2O2,培 養 40 分鐘後,取出 100 μl 菌液系列稀釋至 10-6倍後,均勻塗抹在 LB 固態培養 基上,於 37°C 培養 16 小時後,數菌落數。存活率是根據實驗組每 1 ml 所存活 的菌數,與未加入 3 mM 的 PQ 或 10 mM 的 H2O2的控制組,每 1 ml 的菌數比值; 每次獨立實驗,以三重複數據換算出平均值及標準差,呈現的數據為三次獨立實 驗中具代表性的一次。
2.5.2 紙錠分析(Disc diffusion assay)
細菌在 LB 培養液 37°C 培養 16 小時後,稀釋 40 倍至 LB 液中培養至 OD600 約 0.3~0.4 左右,接著用無菌棉花棒沾取菌液均勻塗抹在 LB 培養基上,接著以 無菌鑷子將紙錠置入培養基中央,再吸取 5 µl 20 mM PQ 或 50 mM H2O2至紙錠 上於 37°C 正置培養隔夜。 2.6 酸性環境壓力生存率測定 細菌在 LB 培養液 37°C 培養 16 小時後,以 20 倍稀釋至 LB,在培養至 OD600 約 0.6~0.7 左右,取 1 ml 菌液至微量離心管,以 15000 rpm 離心 5 分鐘後去掉上 清液,再加入 1 ml 、pH4.4 的 LB 培養液培養 1 小時,使細菌在酸性環境處理前, 有一段時間適應。之後,以 15000 rpm 離心 5 分鐘後去掉上清液,再加入 1 ml 、 pH3.0 的 M9 培養液培養 45 分鐘後,立即取出 100 µl 菌液系列稀釋至 10-6倍後, 均勻塗抹在 LB 固態培養基上,於 37°C 培養 16 小時後,數菌落數。存活率是根 據實驗組每 1 ml、處理 pH3.0 的 M9 培養液所存活的菌數,與僅加入 1 ml、pH4.4 的 LB 的控制組,菌數的比值;每次獨立實驗,以三重複數據換算出平均值及標 準差,呈現的數據為三次獨立實驗中具代表性的一次。
2.7 西方墨點法(Western Blot analysis)
14
鐘後,去掉上清液,再以 200 µl 二次水回溶,並以 95°C、10 分鐘加熱破菌。測 定濃度後,將蛋白依等比例混合蛋白質染劑(0.0626 M Tris-HCl pH6.8、2% SDS、 10 % glycerol、0.01% bromophenol blue 以及 100 mM dithiothreitol),再以 95°C 加熱 10 分鐘,取適量蛋白質(10 μg/lane)加入 13.5% SDS-PAGE 電泳分離蛋白 (80V~100V)。蛋白質經過電泳分離後,將膠上之蛋白質轉漬(400 mA、60 分鐘) 於聚偏二氟乙烯膜(polyvinylidene difluoride, PVDF;Millipore, Billerica, MA, USA) 上,再以溶於 1X TBS(Tris-buffered saline;pH 7.4)的 5%脫脂奶粉 4°C 處理隔夜, 遂以 1X TBS 洗去殘留於膜上的脫脂牛奶。接著,加入一級抗體、室溫 2 小時作 用,以 1X TBS 洗去殘留於膜上非專一性的一級抗體訊號,再以二級抗體(alkaline phosphatase- conjugated anti rabbit immunoglobulin G)在室溫下處理 1 小時,以 1X TBS 洗去殘留於膜上非專一性的二級抗體訊號。隨後,加入鹼性磷酸酶緩衝液 (alkaline phosphatase buffer)、呈色劑 BCIP(5-bromo-4-chloro-3-indolylphosphate)、 NBT(nitro blue tetrazolium chloride)避光呈色。
2.8 蛋白質高量表現之質體建構
高量表現之質體的引子設計,詳列於表三。將欲高量表現蛋白質的編碼區 (coding regions) DNA 片段,以 PCR 增幅後,皆入 yT&A 選殖載體中,再轉殖至 蛋白質表現載體 pETQ31 中,使欲表達蛋白質之 DNA 片段與 pETQ31 黏合,將 質體送入 KpCG43 中,再藉由 0.5 mM 異丙基-β-D-硫代半乳糖苷(Isopropyl-β -D-thiogalactopyranoside, IPTG)誘導,大量表達欲取得之蛋白質。 2.9 蛋白質大量誘導 將帶有欲表達片段的菌株以 LB 培養液 37°C 培養 16 小時後,以 40 倍稀釋 至含有 kanamycin 的 LB 培養液中,培養至 OD600約 0.5~0.6 時,加入 0.5 mM IPTG 誘導目標基因,表現 5 小時後,誘導的細菌以 15000 rpm、3 分鐘離心,去除上 清液後,回溶於二次水中。接著以超音波震盪破菌,收集之蛋白質以 13.5%
15 SDS-PAGE 電泳分離蛋白並確認。 2.10 Pro-Q diamond 染色分析蛋白質的磷酸化 將絲胺酸/蘇胺酸磷酸化激酶(Ser/ Thr Kinase)與欲磷酸化之目標蛋白混合, 並加入緩衝溶液(10 mM ATP、25 mM Tris-HCl、1 mM DTT、5 mM MgCl2、1 mM EDTA、pH7.0)中,於 37°C 水浴槽中反應 30 分鐘,隨後以 95°C 加熱 10 分鐘終 止反應,以 13.5% SDS-PAGE 電泳將蛋白分離。將電泳後的膠,以 Fix buffer(50% 甲醇、10%乙酸)固定蛋白 30 分鐘、重複兩次,以二次水洗淨膠上殘餘的甲醇及 乙酸,隨後以 Pro-Q Diamond 染劑(Invitrogen)染膠 90 分鐘,再用 Destain buffer(Invitrogen)退染三次、每次 30 分鐘,最後以二次水洗淨,並以 UV 光照影。 實驗流程皆參考操作手冊(Invitrogen, Pro-Q Diamond phosphoprotein gel stain)。
2.11 酵母菌凝集測試(Yeast Agglutination)
實驗相關流程參考文獻(Blumer et al., 2005)。細菌在 LB 培養液 37°C 隔夜培
養後,將 1 ml 菌液以 15000 rpm、2 分鐘離心並去除上清液後,分別以 saline (0.85% NaCl)或(溶於 saline 的)5 % D- mannose 回溶,再將此對照組及實驗組混著 1%之 酵母菌(Sigma-Aldrich)於 24 孔盤、反應 5 分鐘,並在室溫下於 100 rpm 的震動 儀(shaker)上晃動,並觀察其結果。 2.12 啟動子(Promoter)分析 2.12.1 啟動子報告載體建構 將預測的啟動子片段或截短片段,以 PCR 增幅後,接入 yT&A 選殖載體中, 再轉殖至啟動子報導質體 placZ15 中,使啟動子片段與 lacZ 報導基因結合,將此 質體以電菌方式送入 Kp∆lacZ 菌株中,再藉由β-半乳糖苷酶(β-galactosidase) 活性評估報告啟動子的活性。
16
2.12.2 啟動子活性分析
根據 Miller 方法(Miller, 1972),細菌在 LB 培養液 37°C 培養 16 小時後,將 菌液 40 倍稀釋至含有抗生素的 LB 培養液中,37°C 培養至 OD600約 0.8~1.0 時, 取 100 μl 菌液於試管中,並混以 900 μl 之 Z buffer (60 mM Na2HPO4, 40 mM NaH2PO4, 10 mM KCl, 1 mM MgSO4, 50 mMβ-mercaptoethanol)、17 μl 之 0.1%SDS、及 35 μl 之氯仿(Chloroform),並混合均勻。在 30°C 水浴槽中反應 10 分鐘後,加入 200 μl 的 4 mg/ml ONPG(ortho-nitrophenyl-β
-D-galactopyranoside)(Sigma-Aldrich),直到反應液呈現黃色,以 500μl 之 1M Na2CO3終止反應,記錄每個反應株 OD420的數值,並以 Miller unit [ 1 Miller unit = 1000* OD420/(t*v* OD600);OD420為加入 ONPG 反應呈黃色後的吸光值;t =反 應時間(min);v =加入反應的體積(ml);OD600 =未反應前菌液的吸光值]換算出 β-半乳糖苷酶活性。每次獨立實驗,以三重複數據換算出平均值及標準差,呈 現的數據為三次獨立實驗中,較具代表性的一次。 2.13 生物膜染色分析 細菌在 LB 培養液中培養 16 小時後,以 200 倍稀釋至 4 ml LB 中,於玻璃 管或加 150 µl 稀釋菌液於塑膠 96 孔盤中,在 37°C 靜置或 50 rpm 搖晃培養 24 或 48 小時,小心去除菌液後,以一次水清洗兩次,再分別加入 4 ml 或 180 µl 1% 結晶紫於玻璃管或塑膠 96 孔盤中,分別以(玻璃管中)靜置或(96 孔盤中)60 rpm 染色處理(玻璃管中)15 或(96 孔盤中)50 分鐘,以一次水清洗黏附不專一的結晶 紫 3 次,96 孔盤再以 95%、180 µl 乙醇溶解結晶紫,並以 ELISA reader 定量。 每次獨立實驗,以五重複數據換算出平均值及標準差,呈現的數據為三次獨立實 驗中具代表性的一次。
17
2.14 剛果紅(Congo red)及鈣螢光(Calcofluor)表現型分析
配置包含 20 µg/mLbrilliant blue、40 µg/mL 剛果紅(Congo red)的(不含 NaCl)LB 固態培養基,及包含 4 mg/L glucose、1mM HEPES、20 µg/L 鈣螢光 (calcofluor)的 LB 固態培養基,將 37°C 培養 16 小時的菌液,各以 10 µl 一次 3 重複滴於剛果紅固態培養基上,在 28°C 培養 48 小時、並於 37°C 再培養 24 小 時,以觀察菌落現象。將 37°C 培養 16 小時的菌液,以接菌環塗佈於鈣螢光固態 培養基上,37°C 避光培養 16 小時後,利用黑燈管照射,以觀察是否有螢光顯現。 呈現的數據為三次獨立實驗中,較具代表性的一次。 2.15 數據分析 氧化壓力或酸性環境生存率測定、啟動子活性分析以及生物膜染色定量分析, 皆為每次獨立實驗,以至少三重複數據換算出平均值及標準差(standard deviation, SD),呈現的數據為三次獨立實驗中具代表性的一次。數據以平均值+標準差呈 現,群組間的差異以雙尾(two tailed)的 Student’s t-test 計算,P-value 小於 0.05 具統計差異。
18
第三章 實驗結果
3.1 K. pneumoniae CG43S3 ∆kpnK_1 的 CPS 生合成
前人研究(萬舉豪,2012)指出,CG43S3 ∆manB 在 4000 rpm 低速離心 5 分 鐘後,比 CG43S3 更容易沉降,顯示 manB 基因缺失會降低 CPS 的生合成(附錄 一 A、B)。若將影響 CPS 前驅物合成的酵素 ManB 之活化位 Ser98 突變成 Alanine, 藉由 Pro-Q Diamond assay 證明,ManB 會失去磷酸化訊號(附錄一 B),且可能因 其酵素活性降低而影響 CPS 的生成(萬舉豪,2012)。如圖一 A、B 所示,前人 建構的 kpnK 基因缺損突變株 kpnK_1,比其親本株 CG43S3 更容易以低速離心沉 降下來,此暗示∆kpnK_1 會影響 CPS 的生合成,但在質體回補株∆kpnK_1 [pRK415-kpnK]卻無法恢復其 CPS 的生成。 3.2 kpnK 下游基因 dsbA 缺失對 CPS 生合成的影響 為了確認此核酸片段移除,是否影響下游基因dsbA的啟動子(圖二A)而造成 polar effect,進而降低CPS的生合成,於是分別建構突變株∆dsbA及∆kpnK∆dsbA, 如圖二B、圖二C,分別將577-bp的dsbA或2021-bp的kpnK與dsbA移除後,以引子 對HY20F/HY21R及HY18F/HY21R進行聚合酶連鎖反應確認,再將突變菌株隔夜 培養後,以4000 rpm低速離心3分鐘,結果發現當∆kpnK_1沉降時,∆dsbA及 ∆kpnK∆dsbA仍呈現與CG43S3相同懸浮的狀態,顯示∆kpnK_1降低了CPS的生成, 並非因為破壞dsbA所導致;而此結果暗示∆kpnK_1突變株可能在kpnK基因外發生 二次突變而影響CPS表現。 3.3 CG43S3 ∆kpnK 基因缺損株的建構與其生長曲線分析 如圖三 A,為了排除前項因素,我重新設計了另一個不破壞 dsbA 啟動子的 kpnK 基因缺失突變體及回補株,分別以引子對 HY18F/HY18R 及 HY19F/HY19R 進行鏈狀聚合反應,產生 504-bp 及 837-bp 的 A、B 片段,並利用 pKAS46 自殺
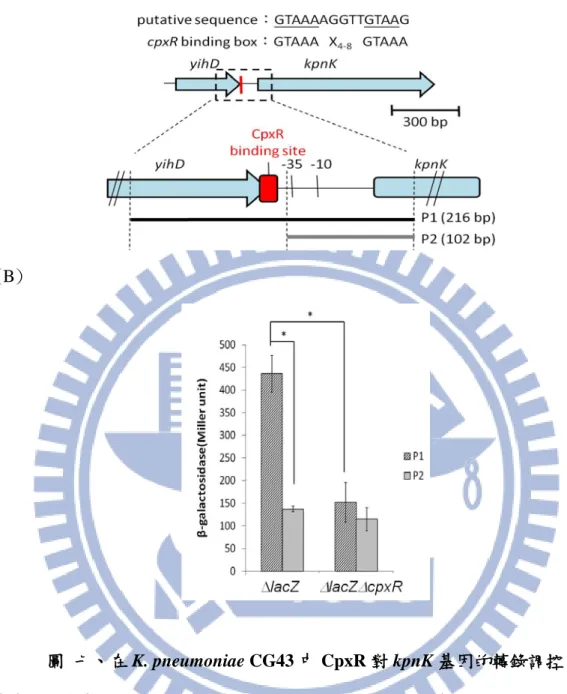
19 性載體承載此 A、B 兩片段,藉兩次同源互換剔除 711-bp 的目標基因後,如圖 三 B,以引子對 HY18F/HY19R 進行鏈聚合酶連鎖反應確認 kpnK 的缺失;如圖 三 C,以引子對 HY18F/HY19R 進行鏈狀聚合酶反應確認 kpnK 回補株。進一步, 透過生長曲線量測結果顯示,不論是在 LB(圖四 A)或是 M9(圖四 B)震盪培養的 條件下,kpnK 基因的缺失對其生長都沒有明顯影響。 3.4 kpnK 基因缺損突變株中 ManB 的磷酸化 圖五 A 將新建構的∆kpnK 及∆kpnK::kpnK 以 4000 rpm 離心 3 分鐘後仍呈懸 浮狀態,此暗示 kpnK 基因缺失並不影響 CPS 的生成;圖五 B 顯示,相對於 CG43S3[pETQ33-ManB],CG43S3[pETQ33-ManB-S98A]中 ManB 位置的磷酸化 訊號會消失,暗示 ManB 受一 STK 所調控,然而∆kpnK [pETQ33-ManB]仍保有 ManB 相對位置的磷酸化訊號,此結果顯示 ManB-S98 磷酸化與 KpnK 可能無 關。 3.5 kpnK 基因缺損株的抗氧化能力分析 . 根據文獻指出在 KP MGH78578 中,當 KpnK 受外界環境壓力刺激後,會影 響細菌的抗氧化能力(Srinivasan et al., 2012)。如圖六 A 所示,將 CG43S3、∆kpnK 及其基因回補株處理 5 mM 的過氧化氫,生存率皆分佈於 75% ~80%間,並無明 顯差異;而圖六 B 指出,在 3 mM paraquate 處理後,CG43S3 和∆kpnK 的生存率 分別為 94%和 64%,且在∆kpnK::kpnK 中也恢復到 84%的生存率,說明∆kpnK 對 paraquate 的氧化壓力有敏感性,但對 H2O2則無差異。 3.6 以 LacZ 報導系統探討 CpxAR 對 kpnK 基因的調控 在大腸桿菌及 Kp MGH78578 中被報導,kpnK 會被具有感受外界壓力的雙 分子調控系統 CpxAR 所影響(Suntharalingam et al., 2003;Srinivasan et al., 2012)。 於是,將 KPCG43S3 的 kpnK 基因上游非轉譯區進行分析,如圖七 A 所示,在啟
20 動子區域含有與 CpxR 鍵結區 GTAAAX4-8GTAAA 極度相似的序列 GTAAAAGGTTGTAAG,定義為可能的 CpxR 鍵結位,於是分別將包含 CpxR 鍵 結區的 P1(216 bp)及不包含 CpxR 鍵結區的 P2(102 bp)片段(圖七 B)與 LacZ 報 導基因結合,再以接合作用轉殖入 KPCG43S3∆lacZ 及 KPCG43S3∆lacZ∆cpxR 菌 株中,分析其β-半乳糖苷酶活性。圖七 C 顯示,失去 CpxR 鍵結區的 P2 活性低 於 P1 啟動子;而 P1 啟動子活性在∆lacZ∆cpxR 中的活性也明顯低於∆lacZ,暗示 kpnK 的表現會受 CpxR 的正向調控。 3.7 kpnK 基因缺損株的抗酸能力 由於 CpxAR 會受諸多的胞外環境壓力,包含:酸鹼度、氧化還原壓力、化 學分子、外膜酯蛋白或膜間質(periplasm)中摺疊錯誤的蛋白所造成的壓力所影響 (Keller and Hunke, 2009; Nakayama and Watanabe, 1995; Snyder et al., 1995),圖七 顯示 KpnK 可能是 CpxAR 調控網路之一員,因此進一步探討 KpnK 和抗酸能力 之相關性。如圖八 A 所示,將 OD600為 0.7-0.8 間對數生長期(log phase)的細菌 培養於 pH4.4 LB 培養液中適應 60 分鐘後放入 pH3.0 M9 培養液中 45 分鐘,再 將培養菌液後系列稀釋後塗盤分析,由 10-5菌落比較發現經酸處理過後,CG43S3 和∆kpnK 及其基因回補株的菌落數差異不大;且由圖八 B 定量菌數顯示,在 CG43S3、∆kpnK 及其基因回補株的生存率分布為 60%~70%間,並沒有因酸處理 後而有明顯差異。 3.8 kpnK 缺損突變株凝集酵母菌的活性分析
在 E. coli 中,CpxAR 會影響線毛的表現,也會正向調控 YdeH,進而抑制細 菌的移動及促進生物膜(biofilm)的生合成(Stefanie et al., 2011)。圖七 B 顯示 CpxAR 在 KPCG43 中正向調控 KpnK 表現,於是,先以酵母菌凝集試驗分析 kpnK 基因剔除對第一型線毛表現的影響。如圖九於 24 孔盤中,將回溶於 0.85%生理 食鹽水的細菌分別在有或無添加 5%甘露醣後,加入 1%之酵母菌,反應結果發
21 現在不表現第一型線毛的親本株 CG43S3 沒有酵母菌凝集反應發生;而∆kpnK 出 現很明顯凝集酵母菌的能力,且此凝集現象會受甘露醣競爭而散去,而 ∆kpnK::kpnK 失去凝集酵母菌的能力,這結果顯示 KpnK 具有負向影響細菌表現 第一型線毛的能力。 3.9 kpnK 基因缺失增加第一型線毛單位蛋白 FimA 的生成 為了探討 KpnK 負向影響第一型線毛單位蛋白 FimA 的表現,如圖十 A 以 FimA 抗體偵測的西方點墨結果得知,kpnK 缺失後 FimA 的表現量會明顯增加, 進一步分析 FimA 啟動子活性發現 kpnK 的缺失也會提高 fimA 啟動子活性(圖十 B),此結果顯示 KpnK 可能透過磷酸化修飾直接或間接影響 FimA 表現。而有關 KP NTUH-K2044 的磷酸化蛋白質體(phosphoproteome)報導指出 FimA 在 Ser、 Thr、Tyr 或 Asp 的殘基上會被磷酸化(Lin et al., 2009,附錄二),而比對 KPCG43 及 KP NTUH-K2044 的 FimA 胺基酸序列發現 T68、T72、S73 及 S74 為可能被磷 酸化的殘基均被保留(圖十 C),FimA 是否被 KpnK 磷酸化還待進一步查證。 3.10 kpnK 基因缺失對第三型線毛表現及生物膜形成能力的影響 第一型及第三型線毛是 KP 最常被報導的胞外膜黏附因子,為探討 KpnK 對 第三型線毛表現的影響,圖十一以西方點墨法分析在 kpnK 基因缺損後 MrkA 的 表現量,發現並無明顯差異。有關克雷白氏肺炎桿菌的研究均指出第三型線毛的 表達與其生物膜的形成有重要的關聯性(Boddicker et al., 2006;Burmolle et al., 2008;Di Martino et al., 2003;Langstraat et al., 2001),如圖十二所示,將隔夜培 養之菌液以 200 倍稀釋,培養 24 小時後並以 1%結晶紫染色觀察及定量其生物膜 的形成,結果發現分別以靜置(圖十二 A)或是搖晃(圖十二 B)培養,不論是在玻 璃管或 96 塑膠孔盤中,CG43S3 及∆kpnK 形成的生物膜皆無明顯差異。
22
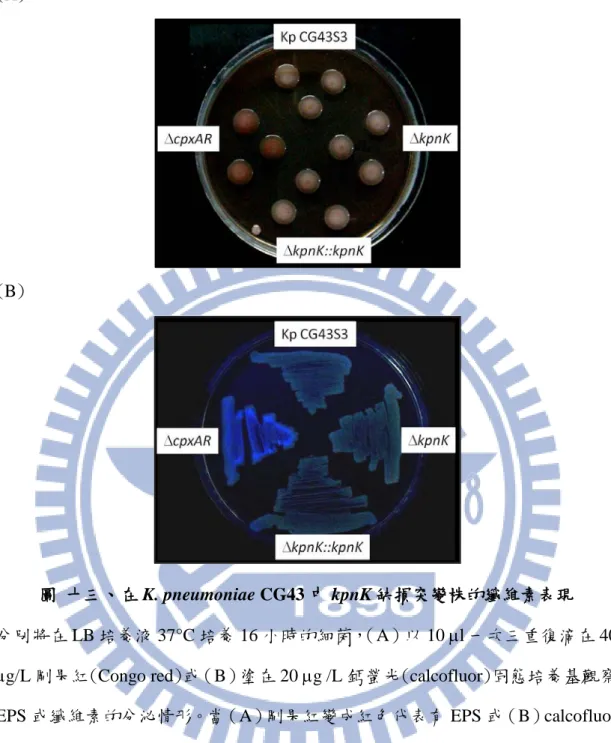
3.11 kpnK 基因缺損株的 EPS 表現
不同的胞外聚合基質(extracellular polymeric substance, EPS)也是決定生物膜 形成的重要因子,在革蘭氏陰性菌中,其組成主要包含纖維素(cellulose)、蛋白 質、核苷酸和酯質。圖十三分別以 40 µg/mL 剛果紅(Congo red)及 20 µg/mL 鈣 螢光(calcofluor)固態培養基觀察細菌 EPS 的分泌情形,當剛果紅染劑和 EPS 鍵 結時,會產生紅色菌落,反之為白色菌落;圖十三 A 中,CG43S3、∆kpnK 及其 基因回補株的 EPS 表現均呈粉色,相對於∆cpxAR 的紅色菌落,均無纖維素的產 生;進一步以對纖維素辨識較專一的鈣螢光固態培養基培養檢測,當纖維素產生 時,利用波長 360nm 的黑燈管照射鈣螢光固態培養基,會有螢光顯現。如圖十 三 B,除了∆cpxAR 外,其他菌株都沒有產生螢光反應,因此推測 KpnK 對纖維 素的生成沒有影響。 3.12 KpnK 定點突變質體的選殖與建構
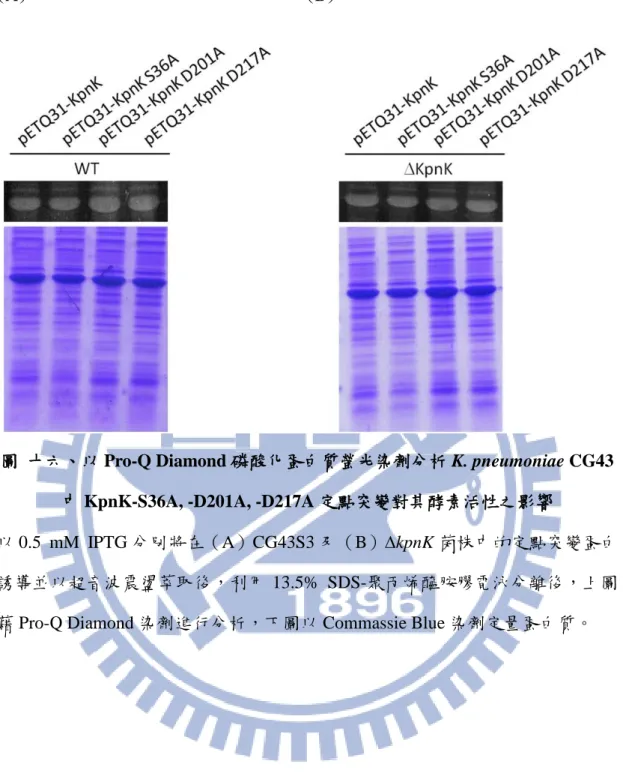
為確認 KpnK 具有激酶活性,由於先前報導指出,E. coli YihE 在 S36、H199、 D201、D217 殘基位置變化會影響其酵素活性(Zheng et al., 2007) 。如圖十四, KpnK 和 YihE 胺基酸序列以 Vector NTI 軟體進行比對,結果發現兩者有高達 80% 的相似度,且在這四個活性殘基均被保留。因此,建構 S36A、D201A、D217A 的 KpnK 突變株,並以 pETQ31 為表現載體(圖十五 A),選殖鏈聚合酶連鎖反應 的產物(圖十五 B) ,並以定序方式確認 S36、D201 及 D217 皆成功突變成 alanine(圖十五 C)。 3.13 KpnK 定點突變對其酵素活性的影響 接著,在細菌中以 IPTG 將這些單點突變的蛋白高量表現,並利用會標示出 Ser/ Thr/ Tyr 被磷酸化的磷酸根染劑 Pro-Q Diamond 來分析 KpnK 激酶活性。結 果顯示,不論是在 CG43S3(圖十六 A)或∆kpnK(圖十六 B)中,
23 變化,暗示著單一活性殘基突變可能不會明顯降低其活性,需幾個活性位點同時 變化才能影響 KpnK 的激酶活性;另一方面,雖然這些點突變使 KpnK 活性降低, 但因為高量表現的結果而無法偵測出其缺失;當然,也無法排除受其他未知激酶 磷酸化的可能性。 3.14 在 CG43S3 中測試 N-及 C-KpnK 的磷酸化 由於 KpnK 的受質未知,而其自我磷酸化的特性曾被報導,為了證實 KpnK 可以本身為受質,首先將可轉譯 kpnK 的 N(766-bp)及 C(241-bp)端的鏈聚合酶 連鎖反應產物分別轉殖至 pETQ31 載體(圖十七 A),再將這些重組質體分別轉型 入 Kp CG43S3 中以 IPTG 誘導蛋白大量表現後,在有無 ATP 的添加處理後,以 電泳膠體展開抽取的總蛋白,分別以 Coomassie Blue 及 Pro-Q Diamond 染色。圖 十七 B 顯示,在未加入 ATP 時,pETQ31-KpnK 和 pETQ31-N-KpnK 即會被磷酸 化,暗示 KpnK 具有自我磷酸化活性或二者皆可受未知激酶磷酸化,圖十七 C 也顯示,pETQ31-C-KpnK 片段蛋白有磷酸化現象,而且,無論 N-KpnK 或 C-KpnK 的磷酸化訊號與是否加入 KpnK 相似。 3.15 N-KpnK 及 C-KpnK 在 kpnK 缺失突變株中激酶活性的探討 為了避免 CG43S3 中內在 KpnK 的活性干擾並探討未知激酶磷酸化 KpnK 的 可能性,將上述質體在∆kpnK 中以 IPTG 誘導蛋白大量表現後(圖十八 A、B), 分別以有無 ATP 處理,藉由電泳膠體展開抽取的總蛋白後,分別以 Coomassie Blue 及 Pro-Q Diamond 染色。圖十八 A 中顯示,在未加入 ATP 時,pETQ31-KpnK 及 pETQ31-N-KpnK 即會被磷酸化,暗示未知激酶參與磷酸化的可能性;圖十八 B 也顯示,pETQ31-C-KpnK 蛋白會受未知激酶磷酸化。
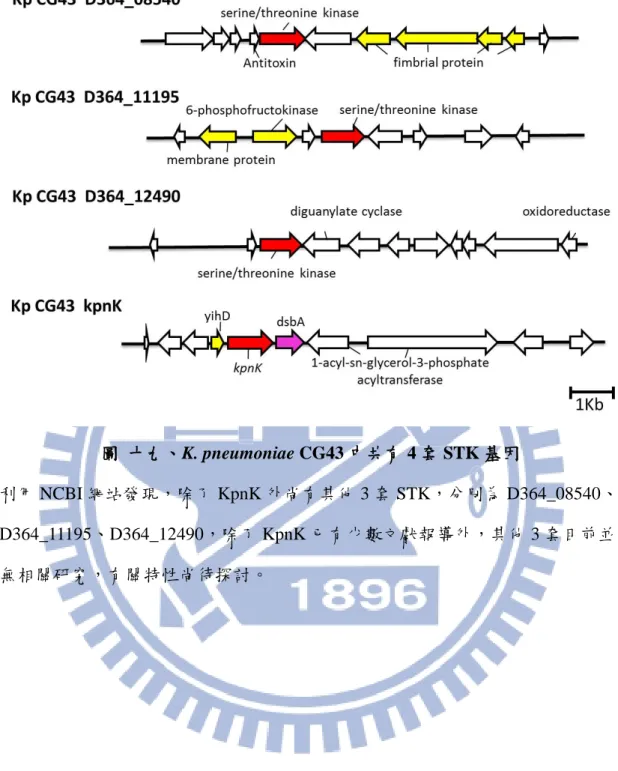
3.16 在 KPCG43 基因體中有多套 STK 基因
24
因體發現,如圖十九所示,除了 KpnK 外尚有 3 套被註解為 STK 的基因,分別 為 D364_08540、D364_11195、D364_12490,然而,除了 KpnK 已有少數文獻報 導外,其他 3 套目前並無相關研究,有關特性尚待探討。
25
第四章 討論
細菌的磷酸化調控路徑中,以雙分子系統最常被報導。但近幾十年間,Ser/ Thr 的磷酸化激酶也在原核生物中被發現,和能量代謝、碳原子降解、胺基酸及 核苷酸的合成、蛋白質運輸以及胞子的形成有關(Li, et al., 2001),雖然 STK 在 真核細胞中已有許多研究,但在原核細菌中的相關研究仍偏少。細菌中,包含原 核型及類真核型的 STK,在 KpCG43 中發現了 KpnK,由於不具完整特定的 ESTK 胺基酸序列,KpnK 為一非典型的 ESTK。 前人研究指出,負責催化 CPS 前驅物的酵素 ManB,在 Ser98 位置會被未知 的激酶磷酸化以影響其酵素活性(萬舉豪,2012;附錄一)。然而,無法以質體回 補其建構的 kpnK_1 基因缺損株的 CPS 表現(圖一)。在S. typhimurium中,yihE-dsbA只會個別轉錄(Suntharalingam et al., 2003),但 E. coli yihE-dsbA 為一基因組 (operon)。且 DsbA 在膜間質(periplasm)具有摺疊錯誤蛋白的能力,因此,推測 DsbA 的缺損可能會增加膜間質壓力,進而造成 CPS 或細菌其他莢膜構造的改變; 為了避免因建構時影響下游基因 dsbA 的表現而間接降低 CPS 的生成,因而重新 建構∆dsbA 及∆kpnK_1∆dsbA 突變菌株,但結果顯示∆kpnK_1 降低 CPS 的生成, 並非因為破壞 dsbA 表現所致。而重新建構不破壞 dsbA 啟動子的 kpnK 基因缺失 突變株,其 CPS 生成和親本株 CG43S3 相似,且 ManB 磷酸化的現象也沒有因 kpnK 的缺失而消失,此結果表示 KpnK 不具磷酸化 ManB-S98 的能力,也驗證 不具影響 CPS 生成的能力。 KpnK 與 YihE 序列相似度達 80%,而 KP CG43 和 KP MGH78578、Kp 342 及 NTUH-K2044 的 KpnK 序列也分別有高達 99%、99%及 80%的序列相似度, 且其 gene organization 也一樣。有趣的是,KPCG43S3∆kpnK 對 H2O2不似 MGH78578∆kpnK 敏感(Srinivasan et al., 2012),可能因為菌株間的差異而造成對 抗 H2O2的能力不同;但在處理 3 mM PQ 後,CG43S3∆kpnK 生存率會降低(圖六 B),顯示其 kpnK 的缺失影響其不同形式之氧化自由基的敏感性。
26
由於在大腸桿菌及 Kp MGH78578 中均報導此 STK 基因會被具有感受外界 壓力的 CpxAR 雙分子系統所調控(Suntharalingam et al., 2003;Srinivasan et al., 2012),而且在 kpnK 啟動子的序列分析中可以發現典型的 CpxR binding box,利 用 LacZ 報導質體分析結果顯示 kpnK 表現受 CpxR 正向調控(圖七 C)。由於 CpxAR 會受諸多的胞外環境壓力,包含酸鹼度壓力,但 CG43S3∆kpnK 對酸的敏 感度與 CG43S3 並無明顯差異。
大腸桿菌 CpxAR 會影響線毛的表現,且沙門氏菌中,KpnK 的同源體 RdoA 會影響纖毛(curli)的生成(Richards., 2006; Zheng et al., 2007)。而實驗結果指出 kpnK 的第一型線毛單位蛋白 FimA 表現和凝聚酵母菌的活性均明顯增加,啟動 子活性分析顯示 kpnK 剔除會增加 fimA 啟動子活性,但由於 KpnK 為一激酶並非 轉錄因子,因此推測可能是透過磷酸化一未知的轉錄因子,進而影響 fimA 的啟 動子活性。在 E. coli 中,fimA 的啟動子會受重組酶 fimB 或 fimE 間接影響,造成 啟動子 Phase-ON 及 Phase-OFF 向位的轉換,進而調控 fim 操縱子的轉錄:當只 有 FimB 坐落於 fimA 啟動子區域時,Phase-ON 及 Phase-OFF 的情況都會發生 (McClain et al., 1991);但當 FimB、FimE 同時坐落此區域時,主要會使得 fimA 啟動子由 Phase-ON 轉換成 Phase-OFF,僅有極少數情況使得 Phase-OFF 轉換成 Phase-ON (Stentebjerg-Olesen et al., 2000)。除此之外,也有直接調控 fimA 啟動 子向位的轉錄因子。H-NS (heat-stable nucleoid-structuring)具有直接鍵結 fimA 啟 動子片段,抑制 fimA 轉錄的能力(Donato et al., 1999; Schembri et al., 1998);或 具有抑制 fimB 和 fimE 的轉錄,間接影響 fimA 向位轉換的能力(Olsen et al., 1994; Olsen et al., 1998; Schwan et al., 2002);或是 H-NS 也會抑制 lrp(leucine-responsive regulatory protein)使得 lrp 無法讓 fimA 啟動子處於 Phase-ON 狀態(Oshima et al., 1995; William, 2013)。且被磷酸化修飾的 H-NS 被發現會影響其鍵結 DNA 的能 力(Arold et al., 2010),雖然 H-NS 目前被發現只在 Tyr 的殘基上被磷酸化,但已 有文獻指出,有些 STK 可能會對 Tyr 的位置進行磷酸化修飾(Ubersax et al., 2007), 因此推測 KpnK 可能透過磷酸化 H-NS 的 Tyr 或其他未知的 Ser/Thr 活化位,而
27
影響 fimA 的啟動子活性(圖二十 B)。有趣的是,KP NTUH-K2044 解出的磷酸蛋 白質體分析發現 FimA 及 FimD 會在特定的 Ser/Thr/Tyr/Asp 被磷酸化(Lin et al., 2009),而這些胺基酸的磷酸化是否與 KpnK 激酶活性相關尚待探討。
此外,在 K. pneumoniae CG43 中,MrkI 會因為將 D56 的位置突變成丙胺酸 (alanine)而降低第三型線毛的表現量,其可能原因是 MrkI 藉由調控 FimB 及 FimE 重組酶的表現,使 fimA 啟動子由 Phase-OFF 轉換成 Phase-ON,而 MrkI 在天門 冬胺酸(aspartate)的磷酸化狀態可能影響其鍵結 fimA 啟動子的能力(吳健誠, 2010),因此,KpnK 不是磷酸化 MrkI D56 的激脢;然而,我們也不能排除 KpnK 磷酸化 MrkI 的 Ser 或 Thr 殘基位置而影響第一型線毛的表現(圖二十 B),但其 相關性仍待探討。
生物膜主要由纖維素、多種菌毛及各種不同的胞外聚合基質(extracellular polymeric substances, EPS)組成。Congo red(CR)用以觀察纖毛(curli)及纖維素的 表現,而 Calcofluor(CF)會和(1-3)-β-或(1-4)-β-D-glucopyranosides 類的纖維素、 chitin 和 succinoglycans 結合(Weiner et al., 1994;Da Re et al., 2006)。在沙門氏菌 中,CF 只會和基因組 bcsABZC 及 bcsEFG 所產生之纖維素產物結合(Solano et al., 2002;Zogaj et al., 2001)。然而,無論 CR 或 CF 分析皆沒能偵測 KpnK 基因缺失 對纖維素合成的影響,此與 KpnK 基因缺失對生物膜合成沒有明顯影響的結果交 互應證。 經 Vector NTI 軟體比對發現 KpnK 保有酵素活性位置 S36、H199、D201、 D217,定點突變結果顯示,KpnK-S36A、-D201A 及-D217A 單一突變可能仍具 有相當程度的激酶活性,因而在 Pro-Q Diamond 染色後,偵辨不出定點突變的影 響,未來可以雙或三定點突變株的建構分析來證實此可能性。除此之外,在 kpnK 的菌株中,KpnK 截短蛋白 N-KpnK 及 C-KpnK 仍有被磷酸化的現象,這個 結果暗示其他未知激酶的影響,而在 CG43 中另外 3 套 STK 是否參與這兩個截 短蛋白及 ManB serine 98 的磷酸化,還有待進一步的研究探討。 本研究指出 K. pneumonia CG43 的 kpnK 在轉錄過程中會受 CpxR 正向調控
28
(圖二十 A);下游會抑制細菌的抗氧化能力及第一型線毛的表現(圖二十 B),而 第一型線毛主要是引起尿道感染的主要原因之一,因此仍須經由動物試驗證實 KpnK 與尿道感染具有直接關聯性。除此之外,未來可設計針對 KpnK 的標的藥 物或是以∆kpnK 菌株誘導而產生的疫苗,以降低細菌所造成的尿道感染。
29
第五章 參考文獻
1. 彭冠男(2013)。克雷白氏肺炎桿菌CG43中RcsFCDB訊息傳遞所扮演的角色。 未出版碩士論文,生物科技所,國立交通大學,新竹市。 2. 樊力成(2012)。YfdX在克雷白氏肺炎桿菌CG43抗酸反應中所扮演的角色。 未出版碩士論文,生物科技所,國立交通大學,新竹市。 3. 林芳瑜(2013)。克雷白氏肺炎桿菌CG43全基因體解序、抗酸轉錄體分析與尿 素酶基因群功能探討。未出版碩士論文,分子醫學研究所,國立清華大學, 新竹市。 4. 林志桓(2009)。RcsB蛋白質在克雷白氏肺炎桿菌CG43中抗酸能力所扮演的角 色。未出版碩士論文,生物醫學所,國立交通大學,新竹市。 5. 黃靜柔(2013)。克雷白氏肺炎桿菌CG43磷酸二酯酶YjcC與二級訊息分子 c-di-GMP在壓力調控之功能性研究。未出版博士論文,分子醫學及生物工程 所,國立交通大學,新竹市。 6. 鄭燕曦(2013)。雙分子系統反應調節因子RcsB在克雷白氏肺炎桿菌CG43中所 扮演的角色。未出版碩士論文,分子醫學及生物工程所,國立交通大學,新 竹市。 7. 鄭威芸(2011)。克雷白氏肺炎桿菌CG43中MrkH、MrkI、MrkJ和CsgD在調控 第一型與第三型線毛表現所扮演的角色。未出版碩士論文,生物科技所,國 立交通大學,新竹市。 8. 曾品瑄(2011)。HdeB、HdeB1、HdeD蛋白質參與克雷白氏肺炎桿菌CG43抗 酸反應的探討。未出版碩士論文,生物醫學所,國立交通大學,新竹市。 9. 吳宛怡(2012)。克雷白氏肺炎桿菌CG43抗氧化逆境反應的研究-超氧化歧化 酶SodA的表現調控。未出版碩士論文,生物科技所,國立交通大學,新竹市。 10. 吳健誠(2010)。克雷白氏肺炎桿菌線毛的分布與表現。未出版博士論文,生 物科技學系,國立交通大學,新竹市。30
11. 萬舉豪(2012)。克雷白氏肺炎桿菌CG43中磷酸甘露糖異構酶磷酸化的特性探 討。未出版碩士論文,生物科技所,國立交通大學,新竹市。
12. Arold, S. T., Leonard, P. G., Parkinson, G. N., & Ladbury, J. E. (2010). H-NS forms a superhelical protein scaffold for DNA condensation. Proc Natl Acad Sci U S A, 107(36), 15728-15732. doi: 10.1073/pnas.1006966107
13. Bakal, C. J., & Davies, J. E. (2000). No longer an exclusive club: eukaryotic signalling domains in bacteria. Trends Cell Biol, 10(1), 32-38.
14. Blumer, C., Kleefeld, A., Lehnen, D., Heintz, M., Dobrindt, U., Nagy,
G.,Michaelis, K., Emody, L., Polen, T., Rachel, R., Wendisch, V. F., Unden, G. (2005). Regulation of type 1 fimbriae synthesis and biofilm formation by the transcriptional regulator LrhA of Escherichia coli. Microbiology, 151(Pt 10), 3287-3298. doi: 10.1099/mic.0.28098-0
15. Boddicker, J. D., Anderson, R. A., Jagnow, J., & Clegg, S. (2006).
Signature-tagged mutagenesis of Klebsiella pneumoniae to identify genes that influence biofilm formation on extracellular matrix material. Infect Immun, 74(8), 4590-4597. doi: 10.1128/IAI.00129-06
16. Boot, I. R., Cash, P., & O'Byrne, C. (2002). Sensing and adapting to acid stress. Antonie Van Leeuwenhoek, 81(1-4), 33-42.
17. Burmolle, M., Bahl, M. I., Jensen, L. B., Sorensen, S. J., & Hansen, L. H. (2008). Type 3 fimbriae, encoded by the conjugative plasmid pOLA52, enhance biofilm formation and transfer frequencies in Enterobacteriaceae strains. Microbiology, 154(Pt 1), 187-195. doi: 10.1099/mic.0.2007/010454-0
18. Canova, M. J., & Molle, V. (2014). Bacterial serine/threonine protein kinases in host-pathogen interactions. J Biol Chem, 289(14), 9473-9479. doi:
10.1074/jbc.R113.529917