I
國立臺東大學生命科學系 碩士論文
Department of Life Science National Taitung University Master Thesis
紅麴菌代謝物 dimerumic acid、deferricoprogen、
monascin 與 ankaflavin 預防酒精性肝損傷之研究
Study on the Prevention of Alcoholic Liver Injury by Monascus -fermented Metabolites Dimerumic acid,
Deferricoprogen, Monascin, and Ankaflavin
指導教授:李俊霖 博士 Advisor: Chun-Lin Lee, Ph.D.
研究生:賴昭如 Student: Chao-Ju Lai
中華民國 104 年 7 月
July , 2015
II
III
I
中文摘要
體內代謝酒精的過程會使肝細胞氧化還原狀態失衡並產生大量自由基,進而 造成肝臟脂質過量堆積與發炎反應,形成肝損傷。紅麴發酵物及其次級代謝物有 許多保健功效,其中紅麴米有研究指出具提升肝臟中抗氧化酵素活性及抑制發炎 細胞激素活性,可保護肝臟免於受到酒精之影響。紅麴黃色素 monascin (MS) 與 ankaflavin (AK) 除可抗發炎外,還具有降體脂與血脂之效用。另外,紅麴菌代謝 產物 dimerumic acid (DMA) 與 deferricoprogen (DFC) 也被證實有良好之抗氧 化作用。但目前尚未釐清紅麴中預防酒精性肝損傷之有效成分。故本研究第一部 分為探討紅麴之抗氧化物質 DMA 及 DFC 於同濃度下改善酒精性肝損傷之潛 力;第二部分則以有效劑量之紅麴黃色素 MS 與 AK 探究其改善酒精性肝損傷 之能力。兩次試驗皆利用 Lieber-DeCarli 流質酒精飼料餵食 C57BL/6J 小鼠六 週,建立酒精性肝損傷之動物實驗模式,試驗期間並同時管餵試驗物質以評估預 防酒精性肝損傷之效果及機制。研究結果顯示,DMA、DFC、MS 與 AK 皆能 降低肝重/體重之比值,且可顯著降低血清中 aspartate aminotransferase (AST)、
alanine aminotransferase (ALT) 及 alkaline phosphatase (ALP) 等肝損傷指標,其中 DFC 和 AK 降低效果較明顯。血清之 triglyceride (TG) 與肝臟中 total cholesterol (TC) 和 TG 均透過四種試驗物而有所降低,又以 MS 和 AK 降低幅度較大,
且組織病理切片結果發現 MS 與 AK 於低劑量下就可預防肝臟中脂質堆積。而 抗氧化試驗結果也指出,DMA、DFC、MS 及 AK 能有效提升抗氧化酵素活性 並降低脂質過氧化程度,以 AK 效果尤佳。以上結果可發現 AK 對於酒精性肝 損傷與脂肪肝有較好之預防效果。對預防酒精性肝損傷機制方面,DMA 與 DFC 乃透過抑制 mitogen-activated protein kinases (MAPK) 家族磷酸化,進一步調降 下 游 發 炎 相 關 蛋 白 tumor necrosis factor-α (TNF-α) 、 interleukin-6 (IL-6) 、 interleukin-1β (IL-1β)、nuclear factor-kappa B (NF-κB)、inducible NOS (iNOS) 與 cyclooxygenase-2 (COX-2),並上調 peroxisome proliferator-activated receptor-γ (PPAR-γ)、nuclear factor-erythroid 2-related factor-2 (Nrf-2) 及 heme oxygenase-1 (HO-1) 之表現量,最終以抑制發炎反應而達到預防酒精性肝損傷的效果;而 MS 與 AK 除可抑制 MAPK 家族磷酸化外,更能直接減少 TNF-α、IL-6 與 IL-1β,
進而降低 NF-κB 及其下游 iNOS 與 COX-2 產生,並增加 PPAR-γ、Nrf-2 與 HO-1,達到預防酒精對肝臟造成損傷。在酒精性脂肪肝部分,DMA、DFC、MS 與 AK 藉由提升 adenosine monophosphate-activating protein kinase (AMPK) 與 peroxisome proliferator-activated receptor-α (PPAR-α),並降低 sterol regulatory element-binding protein-1 (SREBP-1) 與 acetyl-CoA carboxylase (ACC),而這一連 串的調控最終使得 TG 顯著減少,避免酒精性脂肪肝形成。DMA、DFC、MS 與 AK 皆能預防酒精性肝病發生,其中又以 AK 具最佳之預防效果。
關鍵字:酒精性肝損傷、紅麴二次代謝物、dimerumic acid、deferricoprogen、
monascin、ankaflavin
II
Abstract
The process of ethanol metabolism in the body will cause redox reaction of the hepatic cells to lose balance and produce grave amount of free radicals, proceeding to cause hepatic lipid accumulation and inflammation resulting in liver injury. The Monascus-fermented product and its secondary metabolites have many functional effects. Monascus-fermented red mold rice can raise antioxidant enzyme activities in the liver and restrain anti-inflammatory cytokines activities, which can protect the liver against the damage of alcohol. Monascin (MS) and ankaflavin (AK) not only performed anti-inflammatory effect but also had the effect of regulating body fat and blood lipids. Furthermore, dimerumic acid (DMA) and deferricoprogen (DFC) were also proven as the potent antioxidants. However, the functional compounds of Monascus for preventing alcohol liver disease still remains unclear in current.
Therefore, the first part of this research reveals the abilities of reducing alcohol liver disease of Monascus-produced antioxidants DMA and DFC under the same concentration. The second part is to study effect of the effective dose of monascin (MS) and ankaflavin (AK) on the prevention of alcohol liver disease. In both experiments, the C57BL/6J mice were fed Lieber-DeCarli liquid diet for six weeks in order to establish an animal model of alcoholic liver disease. During the stage, the test substances were orally administrated to the ALD mice in order to investigate the prevention effect and the mechanism against ALD.The results state that DMA, DFC, MS and AK all lowered the liver weight/body weight ratio, and visibly decreased liver function indexes such as aspartate aminotransferase (AST), alanine aminotransferase (ALT) and alkaline phosphatase (ALP) of serum, however, DFC and AK had more significant lowering effect. The serum triglyceride (TG) and liver total cholesterol (TC) and TG all decreased through feeding the four test substances, in which MS and AK had more effect. The pathological statin proves that low dosage of MS and AK prevented the lipid deposition in the liver. Furthermore, DMA, DFC, MS and AK effectively increased the antioxidative enzyme activities and lower lipid peroxidation.
However, AK performed the best antioxidative effect. According to these findings, AK is more effective on the prevention of alcoholic liver disease and fatty liver.
Regarding the mechanism of alcoholic liver disease prevention, the DMA and DFC repressed the phosphorylation of mitogen-activated protein kinases (MAPK) family, and further reduced the eypression of the inflammatory-related proteins including tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), interleukin-1β (IL-1β), nuclear factor-kappa B (NF-κB), inducible NOS (iNOS), and cyclooxygenase -2 (COX-2), then increasing the preformance rate of peroxisome proliferator- activated receptor-γ (PPAR-γ), nuclear factor-erythroid 2-related factor-2 (Nrf-2), and heme oxygenase-1
III
(HO-1), resulting in the prevention of alcoholic liver disease development. As for MS and AK, they not only repressed MAPK family phosphorylation but also directly reduced TNF-α, IL-6, and IL-1β levels, and further decreased feedback ativation of NF-κB and the generation of iNOS and COX-2. PPAR-γ, Nrf-2, and HO-1 were also increased, achieving the prevention of alcohol liver disease. Regarding to alchoholic fatty liver, DMA, DFC, MS, and AK raised adenosine monophosphate-activating protein kinase (AMPK) and peroxisome proliferator-activated receptor-α (PPAR-α), and then lowered sterol regulatory element-binding protein-1 (SREBP-1) and acetyl-CoA carboxylase (ACC) levels. Therese regulation finally resulted in the apparent deduction of TG and inhibited the fatty liver development. DMA, DFC, MS, and AK all prevent the development of alcoholic liver disease, however, AK performed the best prevention effect.
keywords: alcoholic liver injury, Monascus-fermented secondary metabolites, dimerumic acid, deferricoprogen, monascin, ankaflavin
IV
縮寫表
AAP (acetaminophen):乙醯胺基酚
ACC (acetyl-CoA carboxylase):乙醯輔酶 A 羧化酶 ADH (alcohol dehydrogenase):酒精脫氫酶
AFL (alcoholic fatty liver):酒精性脂肪肝
AGEs (advanced glycation end products):糖基化終產物 ALD (alcoholic liver disease):酒精性肝臟疾病
ALDH (acetaldehyde dehydrogenase):乙醛脫氫酶 ALP (alkaline phosphatase):鹼性磷酸酶
ALT (alanine aminotransferase):丙氨酸轉胺酶
AMPK (AMP-activating protein kinase):腺苷酸活化蛋白激酶 AST (aspartate aminotransferase):天門冬氨酸轉胺酶
Aβ 1-40 (amyloid β-potein 1-40):類澱粉樣蛋白 1-40 CAT (catalase):過氧化氫酶
COX-2 (cyclooxygenase-2):第二型環氧化酶 CYP2E1 (cytochrome P450 2E1):細胞色素 P450
DMBA (7,12-dimethyl-1,2- benz[a]anthracene):二甲基苯蔥 DOW (deep ocean water):深層海水
ECM (extracellular matrix):細胞外基質
eNOS (endothelial NOS):內皮性一氧化氮合成酶
ERK1/2 (extracellular regulated kinase 1/2):胞外訊息調節激酶 FAS (fatty acid synthase):脂肪酸合成酶
GABA (γ-amino butyric acid):γ-胺基丁酸
GPDH (glycerol-3-phosphate dehydrogenase):甘油-3-磷酸去氫酶 GPx (glutathione peroxidase):穀胱甘肽過氧化酶
GRd (glutathione reductase):穀胱甘肽還原酶 GSH (glutathione):穀胱甘肽
GSH/GSSG (glutathione/ oxidized glutathione):總穀胱甘肽 HCC (hepatocellular carcinoma):肝癌
HDL-C (high-density lipoprotein cholesterol):高密度脂蛋白膽固醇
HMG-CoA (3-hydroxy-3-methylglutaryl CoA):3-羥基-3-甲基戊二酸醯輔酶 HO-1 (heme oxygenase-1):一型血紅素氧化酶
HR-LPL (heparin-releasable lipoprotein lipase):肝素釋放脂蛋白脂解酶 HSCs (hepatic stellate cell):肝星狀細胞
IL-1 (interleukin-1):介白素-1 IL-1β (interleukin-1β):介白素-1β IL-6 (interleukin-6):介白素-6
V
iNOS (inducible NOS):誘發型一氧化氮合成酶 JNK ( Jun N-terminal kinase):c-Jun N 端蛋白質激酶
LDL-C (low-density lipoprotein cholesterol):低密度脂蛋白膽固醇 LPO (lipid peroxidation):脂質過氧化
LPS (lipopolysaccaride):脂多醣
MAPK (mitogen-activated protein kinases):促分裂素原活化蛋白激酶 MDA (malondialdehyde):丙二醛
MEOS (microsomal ethanol oxidizing system):微粒體乙醇氧化系統 MMP-7 (matrix metalloproteinase-7):基質金屬蛋白酶-7
MTP (microsomal triglyceride transfer protein):原漿微粒三酸甘油酯轉移蛋白 NAFLD (non-alcoholic fatty liver disease):非酒精性脂肪肝
nNOS (neuronal NOS):神經一氧化氮合成酶 NO (nitric oxide):一氧化氮
Nrf-2 (nuclear factor-erythroid 2-related factor-2):NF-E2 相關因子-2
PPAR-α (peroxisome proliferator-activated receptor-α):過氧化體增殖活化受體-α PPAR-γ (peroxisome proliferator-activated receptor-γ):過氧化體增殖活化受體-γ RAGE (receptor for advanced glycation endproducts):糖基化終產物受體 ROS (reactive oxygen species):活性氧
SDS-PAGE (sodiumdodecyl Sulfate polyacrylamide gel electrophoresis):十二基硫酸 鈉電泳法
SL (silymarin):水飛薊素
SOD (superoxide dismutase):超氧化物歧化酶
SREBPs (sterol regulatory element-binding proteins):固醇類調節結合蛋白 TC (total cholesterol):總膽固醇
TG (triglyceride):三酸甘油酯
TNF-α (tumor necrosis factor-α):腫瘤壞死因子-α
TPA (12-O-tetradecanoylphorbol-13-acetate):佛波醇-12-十四烷醯-13-乙酸酯 VLDL (very low density lipoprotein):極低密度脂蛋白
WHO (World Health Organization):世界衛生組織
VI
目錄
中文摘要... I Abstract ... II 縮寫表... IV
前言... 1
第一章 文獻回顧... 3
第一節 肝臟疾病 ... 3
一、肝損傷 ... 3
二、肝纖維化 (Hepatic fibrosis) ... 4
三、肝硬化 (Liver cirrhosis) ... 5
四、肝癌 (Hepatocellular carcinoma, HCC) ... 5
第二節、酒精的代謝及酒精性肝損傷 ... 6
一、酒精代謝 ... 6
二、酒精性肝臟疾病 (alcoholic liver disease, ALD) ... 9
三、ALD 之治療方式 ... 15
四、酒精性脂肪肝 (alcoholic fatty liver, AFL) ... 16
第三節 紅麴菌 ... 22
一、紅麴菌之菌種特性 ... 22
二、紅麴菌之分類 ... 24
三、紅麴於預防醫學之應用 ... 24
四、紅麴菌之保健成分 ... 31
五、DMA、DFC、MS 與 AK 之過去研究 ... 35
第二章 研究動機與架構... 41
第一節 研究動機與目的 ... 41
第二節 研究架構 ... 42
第三章 材料與方法... 44
第一節 實驗材料 ... 44
一、實驗儀器與設備 ... 44
二、藥品 ... 44
第二節 動物試驗 ... 45
一、本研究動物試驗模式 ... 45
二、紅麴菌抗氧化物質 DMA、DFC 對酒精性肝損傷之影響 ... 46
三、紅麴菌黃色素 MS、AK 進行預防酒精性肝損傷之動物試驗 ... 49
第四章 研究結果... 57
第一節 紅麴菌抗氧化物質 DMA 與 DFC 對酒精性肝損傷之影響 ... 57
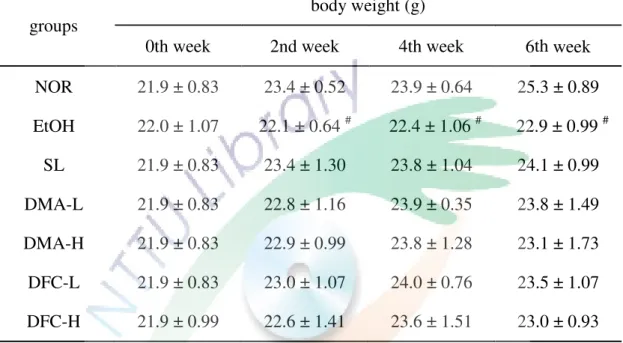
一、DMA 與 DFC 對酒精性肝損傷小鼠體重與肝重/體重比值之影響 .... 57 二、DMA 與 DFC 對酒精性肝損傷小鼠血清中肝功能指標 AST、ALT 及
VII
ALP 含量之變化 ... 57
三、DMA 與 DFC 對酒精性肝損傷小鼠血清與肝臟中脂質濃度之影響 . 57 四、DMA 與 DFC 調節酒精性肝損傷小鼠之脂質過氧化程度 ... 61
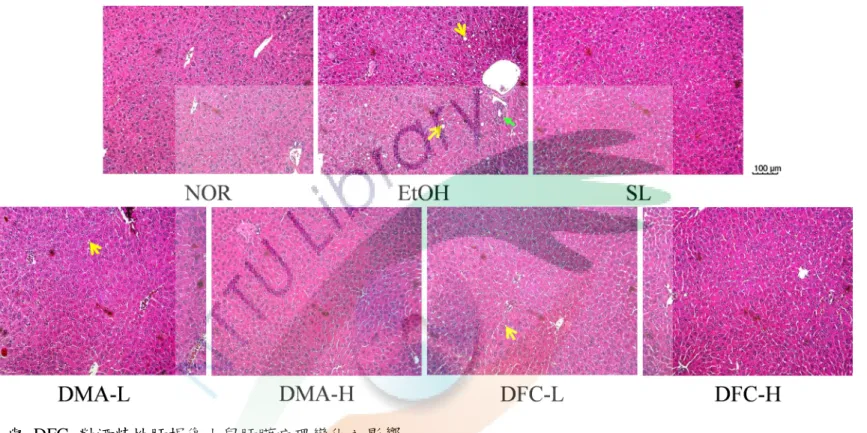
五、DMA 與 DFC 對酒精性肝損傷小鼠肝臟病理變化之影響 ... 61
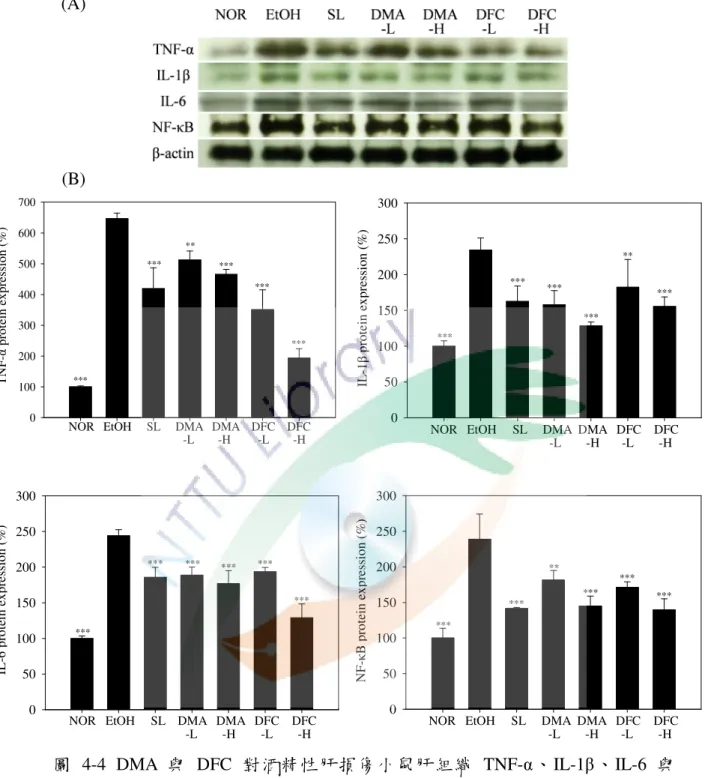
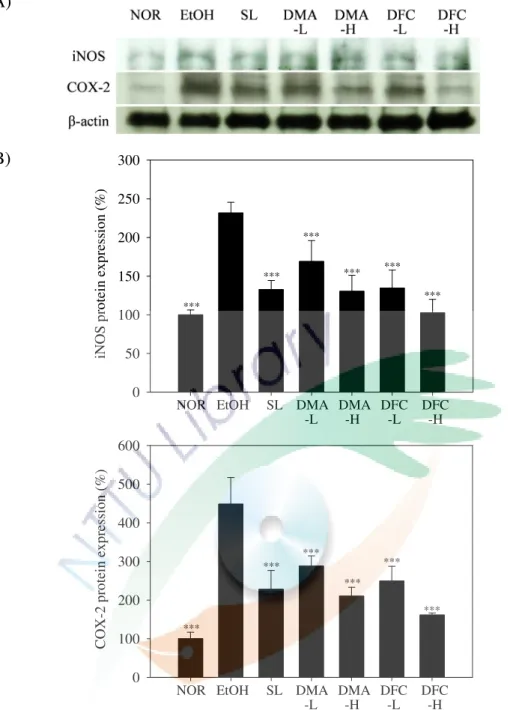
六、DMA 與 DFC 對酒精性肝損傷小鼠肝臟中抗氧化酵素活性之影響 . 61 七、DMA 與 DFC 對酒精性肝損傷小鼠肝臟中發炎相關蛋白表現量之影響 ... 65
八、DMA 與 DFC 對酒精性肝損傷小鼠肝臟中抗氧化發炎蛋白表現量之影 響 ... 71
九、DMA 與 DFC 對酒精性肝損傷小鼠肝臟中脂質代謝與生合成相關蛋白 表現量之影響 ... 74
第二節 紅麴菌黃色素 MS、AK 進行預防酒精性肝損傷之動物試驗 ... 77
一、MS 與 AK 對酒精性肝損傷小鼠體重與肝重/體重比值之影響 ... 77
二、MS 與 AK 對酒精性肝損傷小鼠血清中肝功能指標 AST、ALT 與 ALP 含量之變化 ... 77
三、MS 與 AK 對酒精性肝損傷小鼠血清與肝臟中脂質之影響... 81
四、MS 與 AK 調節酒精性肝損傷小鼠之脂質過氧化程度... 81
五、MS 與 AK 對酒精性肝損傷小鼠肝臟病理變化之影響... 81
六、MS 與 AK 對酒精性肝損傷小鼠肝臟中抗氧化酵素活性之影響... 85
七、MS 與 AK 對酒精性肝損傷小鼠肝臟中發炎相關蛋白表現量之影響 ... 85
八、MS 與 AK 對酒精性肝損傷小鼠肝臟中抗氧化發炎相關蛋白表現量之 影響 ... 89
九、MS 與 AK 對酒精性肝損傷小鼠肝臟中之質代謝與生合成相關蛋白表 現量之影響 ... 89
第五章 討論... 96
一、本研究誘發 C57BL/6J 小鼠酒精性肝損傷之模式 ... 96
二、綜合比較不同試驗物質對酒精性肝損傷小鼠肝功能指標及組織病理之 改善效果 ... 96
三、試驗物質對酒精性肝損傷小鼠之肝臟抗氧化酵素系統之影響 ... 97
四、試驗物質調控酒精誘發肝臟發炎因子表現之能力 ... 98
五、不同試驗物質調控酒精性脂肪肝中脂肪酸代謝與生合成相關蛋白之能 力 ... 99
六、DMA 與 DFC 預防酒精性肝損傷之調控機制 ... 100
七、MS 與 AK 預防酒精性肝損傷之調控機制... 102
第六章 結論... 104
第七章 未來展望... 106
第八章 參考文獻... 107
VIII
圖目錄
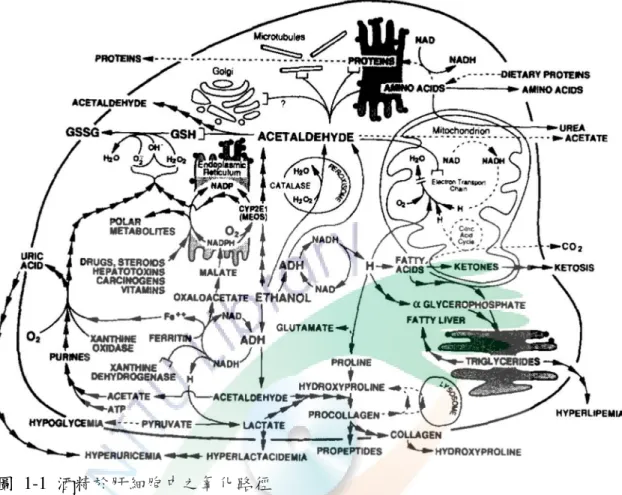
圖 1-1 酒精於肝細胞中之氧化路徑 ... 7
圖 1-2 肝臟中酒精代謝系統 ... 8
圖 1-3 酒精性肝臟疾病之成因 ... 10
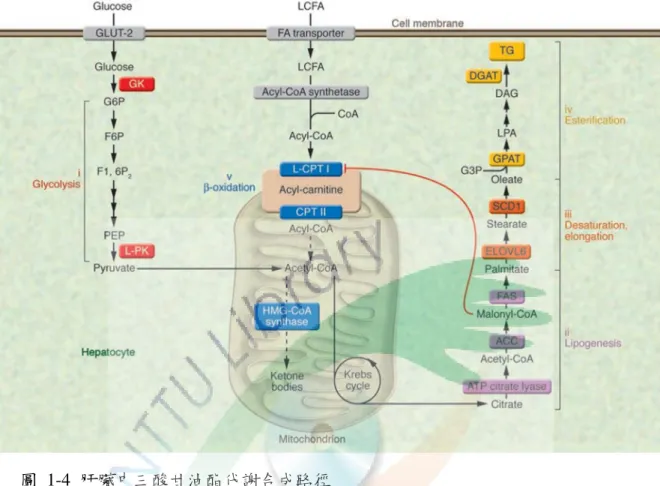
圖 1-4 肝臟中三酸甘油酯代謝合成路徑 ... 17
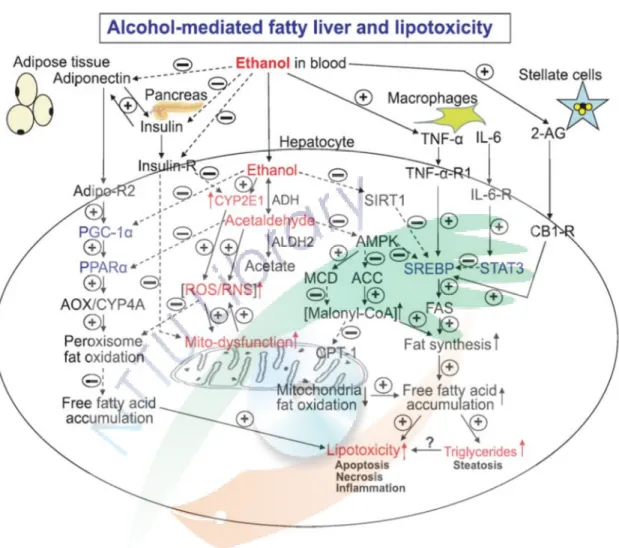
圖 1-5 酒精造成脂肪肝及脂毒性之主要途徑 ... 18
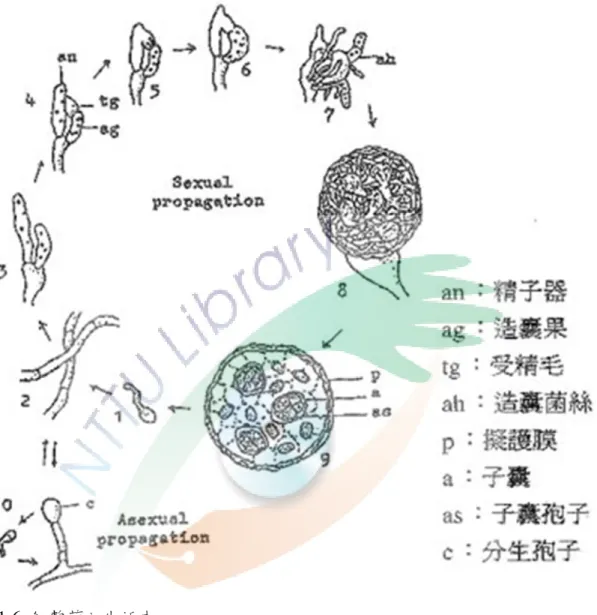
圖 1-6 紅麴菌之生活史 ... 23
圖 2-1 本論文之研究大綱 ... 43
圖 4-1 DMA 與 DFC 對酒精性肝損傷小鼠肝重/體重比值之影響 ... 59
圖 4-2 DMA 與 DFC 對酒精性肝損傷小鼠脂質過氧化之影響 ... 63
圖 4-3 DMA 與 DFC 對酒精性肝損傷小鼠肝臟病理變化之影響 ... 64
圖 4-4 DMA 與 DFC 對酒精性肝損傷小鼠肝組織 TNF-α、IL-1β、IL-6 與 NF-κB 之影響 ... 67
圖 4-5 DMA 與 DFC 對酒精性肝損傷小鼠肝組織 iNOS 和 COX-2 之影響 . 69 圖 4-6 DMA 與 DFC 對酒精性肝損傷小鼠肝組織 ERK1/2 和 p-ERK1/2 之影 響... 70
圖 4-7 DMA 與 DFC 對酒精性肝損傷小鼠肝組織 p38 和 p-p38 之影響 ... 72
圖 4-8 DMA 與 DFC 對酒精性肝損傷小鼠肝組織 PPAR-γ、Nrf-2 和 HO-1 之 影響... 73
圖 4-9 DMA 與 DFC 對酒精性肝損傷小鼠肝組織 AMPK、p-AMPK 和 PPAR-α 之影響... 75
圖 4-10 DMA 與 DFC 對酒精性肝損傷小鼠肝組織 SREBP-1 和 ACC 之影響 ... 76
圖 4-11 MS 與 AK 對酒精性肝損傷小鼠肝重/體重比值之影響 ... 79
圖 4-12 MS 與 AK 對酒精性肝損傷小鼠脂質過氧化之影響 ... 83
圖 4-13 MS 與 AK 對酒精性肝損傷小鼠肝臟病理變化之影響 ... 84
圖 4-14 MS 與 AK 對酒精性肝損傷小鼠肝組織 TNF-α、IL-1β、IL-6 與 NF-κB 之影響... 87
圖 4-15 MS 與 AK 對酒精性肝損傷小鼠肝組織 iNOS 和 COX-2 之影響 ... 88
圖 4-16 MS 與 AK 對酒精性肝損傷小鼠肝組織 ERK1/2 和 p-ERK1/2 之影響 ... 90
圖 4-17 MS 與 AK 對酒精性肝損傷小鼠肝組織 p38 和 p-p38 之影響 ... 91
圖 4-18 MS 與 AK 對酒精性肝損傷小鼠肝組織 PPAR-γ、Nrf-2 和 HO-1 之影 響... 92
圖 4-19 MS 與 AK 對酒精性肝損傷小鼠肝組織 AMPK、p-AMPK 和 PPAR-α 之影響... 94
圖 4-20 MS 與 AK 對酒精性肝損傷小鼠肝組織 SREBP-1 和 ACC 之影響 .... 95
IX
圖 4-21 DMA 與 DFC 預防酒精性肝損傷之相關調控機制 ... 101 圖 4-22 MS 與 AK 預防酒精性肝損傷之相關調控機制 ... 103
X
表目錄
表 1-1 紅麴發酵之代謝物於預防醫學之應用 ... 32 表 3-1 試驗飼料組成 ... 47 表 3-2 紅麴菌次級代謝物 DMA 與 DFC 對預防酒精誘發小鼠肝損傷試驗之動
物分組與試驗物質劑量配置 ... 48 表 3-3 紅麴菌次級代謝物 MS 與 AK 對預防酒精誘發小鼠肝損傷試驗之動物
分組與試驗物質劑量配置 ... 50 表 4-1 DMA 與 DFC 對酒精性肝損傷小鼠體重之影響 ... 58 表 4-2 DMA 與 DFC 對酒精性肝損傷小鼠血清肝功能指標 AST、ALT 與 ALP 含量之影響... 60 表 4-3 DMA 與 DFC 對酒精性肝損傷小鼠血清與肝臟中膽固醇及三酸甘油酯
之影響... 62 表 4-4 DMA 與 DFC 對酒精性肝損傷小鼠肝臟中抗氧化酵素活性之影響 ... 66 表 4-5 MS 與 AK 對酒精性肝損傷小鼠體重之影響 ... 78 表 4-6 MS 與 AK 對酒精性肝損傷小鼠血清肝功能指標 AST、ALT 與 ALP 含
量之影響... 80 表 4-7 MS 與 AK 對酒精性肝損傷小鼠血清與肝臟中膽固醇及三酸甘油酯之影
響... 82 表 4-8 MS 與 AK 對酒精性肝損傷小鼠肝臟中抗氧化酵素活性之影響 ... 86 表 4-9 DMA、DFC、MS 與 AK 預防酒精性肝損傷效果... 105
1
前言
世界衛生組織 (World Health Organization, WHO) 之全球肝癌患者統計報告 顯示,全球肝癌患者約 71% 集中於亞洲地區。衛福部所公告之 102 年國人十 大死因中,慢性肝病及肝硬化排名第九,而位居首位的癌症裡,肝癌為第二位 (行 政院衛生福利部,2014)。飲用酒精性飲料於現今社會越來越普遍,雖有許多研 究指出適量飲酒能有助於減少心血管疾病的發生 (Moore and Pearson, 1986),但 過量會使肝臟無法立即代謝,造成損傷。根據衛福部統計,我國有 54.6% 民眾 表示過去曾經喝酒,在喝過酒的民眾裡有 47.1% 曾有喝醉之經驗,表示國人罹 患酒精性肝臟疾病的風險不低。肝臟疾病初期為肝損傷,其中慢性肝損傷會導致 肝臟走向纖維化、肝功能衰竭與肝癌 (Malhi and Gores, 2008),其誘因有脂肪肝、
病毒性、酒精性等,並會互相影響加速肝臟損傷。肝臟中酒精代謝於有三條路徑,
分 別 為 酒 精 脫 氫 酶 (alcohol dehydrogenase, ADH) 、 微 粒 體 乙 醇 氧 化 系 統 (microsomal ethanol-oxidizing system, MEOS) 之細胞色素 P450 (cytochrome P450 2E1, CYP2E1) 和 catalase (CAT) (Lumeng and Crabb, 2000; Seitz and Stickel, 2007),而在代謝過程中造成肝細胞氧化還原狀態不平衡以及產生之自由基為近 幾年主要探討酒精性肝損傷之方向 (Lieber, 1985; Porta, 1997)。另外,酒精會影 響脂肪合成及降解平衡之關鍵產物 (You et al., 2004) 及誘導脂質合成酵素 (Sozio and Crabb, 2008),造成酒精性脂肪肝。
紅麴菌 (Monascus spp.) 應用於飲食及醫藥已有好幾世紀,更於近年來逐漸 發展為保健食品。先前研究中紅麴發酵物之效用,包含可提升小鼠肝臟中抗氧化 酵素之活性及抑制發炎相關因子之表現量,降低酒精對肝細胞所造成之損傷 (Cheng and Pan, 2011);紅麴山藥能夠抗動脈粥狀硬化 (Lee et al., 2007a);紅麴發 酵之大豆萃取物與紅麴發酵混合榖物萃取物可降低高血脂症 (Pyo and Seong, 2009; Pyo and Seong, 2013);紅麴發酵黑豆抑制體脂肪形成 (Oh et al., 2014);紅 麴薏仁則具調節血糖之功效 (Shi and Pan, 2010a) 等。而近年來則是以紅麴次級 代謝物為研究方向,進一步探討功效成分,其中又以黃色素 monascin (MS) 與 ankaflavin (AK) 最被廣泛研究。文獻指出,MS 與 AK 皆可透過減少發炎因子 達到抗發炎效果 (Hsu et al., 2011a; Hsu et al., 2012d),此外,兩者亦可透過調控 Nrf-2 產生抗發炎與抗氧化等作用,提高胰島素敏感性,預防如第二型糖尿病等 疾病所造成之發炎反應 (Lee et al., 2012; Lee et al., 2013c) 等。而除了紅麴色素外,
2
也有越來越多學者探討其他功效成分,如 dimerumic acid (DMA),具清除自由基 之能力,降低 ROS 含量 (Aniya et al., 2000),並可調控 Nrf-2 抑制發炎細胞激 素 (Lee et al., 2013a) ,同時也具有抗 纖維 化之效用 (Lee et al., 2013b) 。 deferricoprogen (DFC) 則是具良好之抗氧化活性 (Hsu et al., 2012b),且有護腎作 用 (Hsu et al., 2012b)。雖已有研究證實紅麴米可改善酒精性肝損傷,且 DMA 與 DFC 良好的抗氧化效果、MS 及 AK 具抗發炎能力也被許多研究指出,但尚未 有研究探討其預防酒精性肝損傷之功效成分以及相關路徑。
本研究以 silymarin 做為正控制組,其目前在肝臟疾病上之應用機制包括抗 發炎、抗氧化、抗纖維化與免疫調節 (Lieber et al., 2003),試驗物質則為紅麴抗 氧化成分 DMA 和 DFC,以及目前具多種功效之抗發炎物質 MS 和 AK 為主,
並 分 別 探 討 個 別 對 於 酒 精 性 肝 損 傷 之 改 善 能 力 。 兩 次 動 物 試 驗 皆 以 Lieber-DeCarli nutrient diet 長期餵食 C57BL/6J 雄性小鼠來誘發酒精性肝損傷,
預養 2 週後,給予液態飼料任食,並同時管餵不同濃度之試驗物質,試驗 6 週 後觀察其對血清肝功能指標與 TC、TG 之含量影響,再以組織病理切片染色評 估肝臟中脂質堆積程度,也透過西方墨點法了解肝組織中 IL-6、IL-1β、TNF-α、
NF-κB、iNOS、COX-2、ERK1/2 和 p38 等發炎細胞激素與調控發炎因子,及 抗發炎因子 Nrf-2、HO-1 與 PPAR-γ,此外,也分析脂質代謝與生合成相關因 子 AMPK、SREBP-1、ACC 和 PPAR- α 蛋白表現,以探討 DMA 和 DFC 以 及 MS 和 AK 預防酒精性肝損傷之可能機制。
3
第一章 文獻回顧
第一節 肝臟疾病根據行政院衛福部公告 102 年國人十大死因資料顯示,慢性肝病及肝硬化 佔居第九位,而其中肝癌又為所有癌症中之第二位,由此可知肝臟疾病為台灣主 要疾病之一 (行政院衛福部,2014)。肝臟疾病可分為三大階段,即肝損傷 (脂肪 肝與肝炎)、肝纖維化/肝硬化以及最後一個階段肝癌,其發生原因又分為以下幾 點:
一、肝損傷
肝損傷分為急性與慢性,而其中慢性肝損傷又是使肝臟走向纖維化、肝功能 衰竭與肝癌的主要原因 (Malhi and Gores, 2008),其定義為血清丙氨酸轉胺酶 (alanine aminotransferase, ALT) 或結合型的膽紅素高於正常值上限的兩倍,或同 時合併有血清天門冬氨酸轉胺酶 (aspartate aminotransferase, AST)、鹼性磷酸酶 (alkaline phosphatase, ALP) 及總膽紅素增加,任一項高於正常值上限的兩倍 (Navarro and Senior, 2006)。
造成肝損傷有很多種因素,而肝臟也會產生不同的修復機制,分泌細胞激素、
生長因子及其他傳遞因子等 (Michalopoulos and DeFrances, 1997),協助肝臟恢復 原本狀態並重建功能 (Fausto et al., 2006)。肝損傷包括相關肝臟疾病和組織損傷,
而肝損傷的發生原因大致分為以下幾種:
(一) 化學性肝損傷
即化學物質所造成之肝損傷。人類在自然環境中接觸大量外來異物是無可避 免的,例如:化學試劑、人工合成食品添加劑、工業汙染物、殺蟲劑或農藥等,
而這些外來異物可經由各種途徑進入人體造成危害,包括攝入、呼吸及皮膚接觸 (Einolf et al., 1997; Kari et al., 1999)。其中又以四氯化碳 (carbon tetrachloride,
CCl4) 最常被用來作為誘發化學性肝損傷之模式,其原理為肝臟中之微粒酵素活
化四氯化碳後形成三氯甲烷自由基,而後與蛋白質結合導致蛋白質合成受阻,並 造成脂質分解代謝失調,使三酸甘油酯蓄積於肝細胞中。除此之外,三氯甲烷自 由基也會形成過氧化物,促使脂質過氧化造成肝細胞損傷,最終導致肝中酵素滲 出及細胞壞死 (Chen et al., 2012)。
4 (二) 酒精性肝損傷
指長期酗酒所導致之肝損傷。酒精會經由肝臟酵素 CYP2E1 進行代謝,產 生 ROS 造成脂質過氧化、破壞 DNA 及使酵素失活進而引發細胞毒性,使肝細 胞凋亡及壞死 (Zakhari, 2006);此外,酒精也會損害粒線體氧化功能,耗損還原 態穀胱甘肽 (glutathione, GSH),降低肝臟之自我保護功能 (Zakhari, 2013)。
(三) 病毒性肝損傷
病毒引起之肝炎、肝硬化及肝癌。目前已知病毒性肝炎病毒共有八種,分別 為 A、B、C、D、E、G、TT 及 SEN 型肝炎病毒,其中 A 型與 E 型經由糞 口傳染,不常發生慢性肝炎之現象;B、C、D、G 型等則是透過血液或體液之 接觸傳染,最常見為 B 型與 C 型病毒,主要引起慢性肝病、肝硬化、肝癌及 末期肝衰竭;D 型病毒為一種類病毒,需透過 B 型病毒之抗原才可進行複製;
G、TT 與 SEN 型病毒於近十年才被發現,因此對人類肝臟疾病之機轉仍有待 探討與研究 (Poovorawan et al., 2002)。
(四) 藥物性肝損傷
泛指因服用藥物而對肝臟有毒害作用所造成之肝病,目前以非類固醇類消炎 止痛藥例如普拿疼成分乙醯胺基苯酚 (acetaminophen, AAP) (Leonis et al., 2013)、
抗感染劑如 terbinafine (van 't Wout et al., 1994) 及中樞神經用藥為最常見之三大 類藥物 (Abboud and Kaplowitz, 2007; Lucena et al., 2008)。藥物性肝損傷分為可 預期型及不可預期型,可預期型肝損傷與藥物劑量相關,為藥物或其代謝物直接 作用,而大部分藥物則是屬不可預期型肝損傷,其發生原因與免疫反應有關 (Abboud and Kaplowitz, 2007)。
(五) 脂肪肝
脂肪肝是指肝脂肪含量超過全肝臟重量的百分之五,或是在肝組織切片中超 過百分之十以上的肝細胞呈現脂肪空泡變性的現象而稱之 (Brunt, 2001),其中最 常見病因為肥胖、酗酒、糖尿病控制不佳、血脂肪過高、體重過重等因素造成脂 肪在肝細胞中積聚 (Bedogni and Bellentani, 2004)。
二、肝纖維化 (Hepatic fibrosis)
5
肝纖維化為肝損傷所共有的病理變化,是一種持續性傷口癒合反應,使肝 臟纖維組織異常增生,而隨著纖維的產生,影響肝臟中血液的流動,進而造成門 脈壓升高,導致肝功能失調 (Li and Friedman, 1999)。當肝臟受到酒精、病毒、
藥物、金屬離子沈積、自體免疫反應等因素引起肝細胞受損、凋亡或是壞死時,
會使肝臟中之免疫細胞 kupffer cell 活化並分泌細胞激素,刺激竇狀隙中休止狀 態之肝星狀細胞 (hepatic stellate cell, HSC) 活化,進而產生以膠原為主的細胞外 基質 (extracellular matrix, ECM) 並沉積在竇狀隙中,當膠原纖維過度累積於肝 臟中便會導致纖維化 (Li and Friedman, 1999; Stickel et al., 2002)。肝纖維化屬可 逆過程,若能移除致病因子,肝臟本身能改善纖維化之情形,但若肝臟受損形況 持續便會造成肝小葉結構改變,形成假小葉和結節,此階段即為肝硬化,屬不可 逆過程,此時大量的細胞外基質圍繞於肝細胞周圍,長期傷害會造成靜脈曲張、
腹水等併發症,並極有可能導致肝衰竭及肝癌 (Lebrec, 1994)。
三、肝硬化 (Liver cirrhosis)
肝硬化為肝纖維化之晚期病變,是一種慢性肝臟疾病,由許多因素所引起的 肝細胞病變、壞死並伴隨各種發炎反應,當肝臟嚴重受損時,細胞便以更多胞外 基質加以修補,長期下來肝臟便會嚴重纖維化、結構破壞扭曲及形成結節,進而 衍生出肝硬化 (Lebrec, 1994)。肝硬化會導致門脈高壓、凝血病變、血小板減少、
腹水、水腫、黃疸、肝腦病變和肝腎症候群等併發症 (American Gastroenterological, 2002),同時肝硬化的病人,也是發生肝癌的高危險群 (Anselmi et al., 2002)。
四、肝癌 (Hepatocellular carcinoma, HCC)
肝癌在全球常見癌症排名中位於第五位,每年約有 100 萬人確定罹患肝癌,
並會有 50 至 100 萬人會因肝癌而死亡,若按區域性統計,全球肝癌患者約 80%
集中於發展中國家,其中又以東南亞與非洲居多 (Parkin et al., 1984)。根據 2004 年 WHO 全球肝癌患者統計結果,東南亞地區罹患肝癌人數約 6.4 萬人,且西 太平洋海域國家人數更是高達 38.6 萬人,顯示出全球肝癌患者約 71% 集中於 亞洲地區。衛福部 100 年癌症登記年報指出當年初次診斷罹患肝癌者共計 11,292 人,而當年共 8,022 人因肝癌身亡,由此可推估台灣罹患肝癌人數以每 年約一萬人的速度在成長。
肝癌本身為一種多元誘因之疾病,其相關危險因子主要是、B 型肝炎、C 型
6
肝炎、肝硬化,而外在原因為黃麴毒素、飲酒過量、香菸中的尼古丁、嚼食檳榔,
其他本身之因素例如肥胖、非酒精性脂肪肝及糖尿病等皆會導致肝癌發生 (Bosch et al., 1999)。肝癌若依病理分類可分為四種主要類型,分別為肝細胞癌 (hepatocellular carcinoma, HCC),為最常見之肝癌種類,在成人肝癌中約佔 85%;
膽管癌 (cholangiocarcinoma) 發生於組成膽管的細胞,約佔肝癌的 12%;血管肉 瘤 (angiosarcoma) 又稱血管瘤 (hemangiosarcoma),是一種極端稀少的癌症,只 佔肝癌之 1%;最後為肝母細胞瘤 (hepatoblastoma) 通常好發於兒童,是非常罕 見之肝癌種類 (Chung et al., 2009; Kumar et al., 2011)。
第二節、酒精的代謝及酒精性肝損傷
飲用酒精性飲料於現今社會已是越來越普遍的現象,許多研究指出適量飲酒 能有助於減少心血管疾病的發生 (Moore and Pearson, 1986),但過量飲酒則是會 導致身體損傷。酒精主要是由胃部及小腸吸收,但少部分酒精也會透過口腔及大 腸吸收至體內,而酒精除了會使組織中之蛋白質凝固之外,也會與水形成小分子,
不與血漿中蛋白質產生反應,具極佳穿透性,因此可經由血液迅速流動至各個組 織中 (Svensson et al., 1999)。作用後之酒精於人體內會影響中樞神經系統,使飲 用者產生視覺及聽覺障礙,進而降低注意力及判斷力等,其影響程度與體內酒精 濃度成正比 (凌與陳,2000)。
一、酒精代謝
由於酒精無法儲存於人體中,因此有 95% 的酒精必須透過肝臟代謝為水與 二氧化碳排出。圖 1-1 所示,當酒精進入肝臟後會透過酒精脫氫酶 (alcohol dehydrogenase, ADH)、微粒體乙醇氧化系統 (microsomal ethanol-oxidizing system, MEOS) 之 CYP2E1 和 CAT 這三條途徑氧化為乙醛 (Lumeng and Crabb, 2000;
Seitz and Stickel, 2007),乙醛又會被乙醛脫氫酶 (acetaldehyde dehydrogenase, ALDH) 代謝成醋酸,而醋酸經由乙醯輔酶 A 合成酶 (Acetyl-CoA synthetase) 轉變成乙醯輔酶 A,以利肝以外之組織例如心臟等利用 (Lieber, 2005b; Yamamoto et al., 2011)。酒精於肝臟中透過各種酵素系統代謝之詳細反應如圖 1-2:
7 圖 1-1 酒精於肝細胞中之氧化路徑
Fig. 1-1 Oxidation of ethanol in the hepatocyte (Lieber, 1997)
8
圖 1-2 肝臟中酒精代謝系統
Fig. 1-2 Alcohol metabolism in liver (Lieber, 2005b; Yamamoto et al., 2011)
9
(一) 酒精脫氫酶 (alcohol dehydrogenase, ADH)
位於細胞質中,當血液中酒精濃度較低時,主要是透過此酵素作用進行代謝。
此酵素特點為其代謝速率受 NAD+ 影響,因酒精代謝為乙醛過程中會產生
NADH,而 NADH 必須氧化為 NAD+ 才能使酒精繼續代謝,因此 NAD+ 為此 反應速率之限制因子。
(二) 微粒體乙醇氧化系統 (microsomal ethanol oxidizing system, MEOS)
MEOS 位於內質網中,研究發現體內之 MEOS 系統是透過誘導 CYP2E1 來代謝酒精轉化為乙醛,不同的是此代謝方式會產生大量 ROS,其中又以超氧 自由基 (superoxide free radicals) 與氫氧自由基 (hydroxyl free radicals) 為最多 (Porta, 1997),且研究也指出此酵素系統為長期飲酒者主要之代謝途徑,最多可 代謝人體酒精總量約 40% (Lands, 1998)。
(三) 過氧化氫酶 (catalase, CAT)
存在於過氧化小體 (peroxisome) 內,其代謝酒精之方式主要為透過催化過 氧化氫代謝成水,因此 CAT 代謝速率與過氧化氫含量有關,雖然當血液中酒精 濃度超過 0.02% 時 CAT 就會產生作用,不過因體內自然產生過氧化氫之效率 相當低,所以此一途徑可代謝酒精之能力非常有限 (Lands, 1998; Lieber, 1982)。
二、酒精性肝臟疾病 (alcoholic liver disease, ALD)
ALD 之成因如圖 1-3 所示。ALD 是指長期過量飲酒導致之肝臟損傷與病 變,其病理發展與發生率包含酒精性脂肪肝 (alcoholic fatty liver;80-90%)、酒 精性肝炎 (alcoholic hepatitis;10-35%)、酒精性肝纖維化 ( liver fibrosis;20-40%)、
酒 精 性 肝 硬 化 (cirrhosis ;8-20%) 及肝癌 (hepatocellular carcinoma ;3-10%) (Altamirano and Bataller, 2011; McCullough and O'Connor, 1998)。早期對於 ALD 之研究大多以酒精代謝產物乙醛為主要研究焦點,大量乙醛可與細胞膜蛋白及血 中蛋白質結合,影響許多器官正常代謝 (Lin et al., 1988),例如白蛋白、脂蛋白、
粒線體蛋白質及游離胺基酸等 (Donohue et al., 1983; Nomura and Lieber, 1981;
Stevens et al., 1981),而這也是攝入大量酒精後會有噁心、嘔吐、頭痛及血壓上 升等宿醉現象之原因 (Lieber, 1994)。而近年來則是認為 ALD 之形成與肝細胞
10
圖 1-3 酒精性肝臟疾病之成因
Fig. 1-3 The pathogenesis of alcoholic liver disease (Tilg and Day, 2007)
11
氧化還原狀態不平衡以及代謝過程所產生之自由基有關 (Lieber, 1985; Porta, 1997)。
肝細胞中氧化還原狀態不平衡係指 NADH/NAD+ 比值增加,因 ADH 及 ALDH 參與酒精代謝過程中會大量消耗 NAD+,同時產生大量的 NADH,而此 現象會干擾肝細胞之正常代謝功能,進而抑制脂肪酸氧化、促進脂質合成,造成 脂質堆積形成脂肪肝 (Zakhari, 2006),此外,NADH 增加也會催化丙酮酸合成 乳酸,導致血中尿酸濃度過高產生尿酸結晶,最後使痛風發作 (Klatsky et al., 1986)。慢性酒精攝取也會使體內 MEOS 系統誘導 CYP2E1 代謝酒精,產生大 量 ROS,而 ROS 會破壞肝細胞中之 DNA、蛋白質與脂質等重要物質,使 DNA 斷裂、酵素無法活化、蛋白質變性、脂質過氧化,造成肝臟受損 (Lieber, 2005b)。
另外,酒精也會降低體內抗氧化酵素活性及抗氧化能力,例如超氧化物歧化酶 (superoxide dismutase, SOD)、穀胱甘肽過氧化酶 (glutathione peroxidase, GPX) 等,
促使過氧化脂質大量產生,引起 ALD (Rouach et al., 1997)。除此之外,代謝產 物乙醛亦會造成肝臟中抗氧化劑如穀胱甘肽 (glutathione, GSH) 含量大幅降低 (Oh et al., 1998)。
除了間接造成肝損傷之外,酒精也會直接影響肝細胞之生長週期,Wands 等人之研究建立肝損傷模式後再進行肝臟局部切除手術,發現若於術後繼續攝入 酒精,其肝細胞中 DNA 複製及合成能力大幅降低;反之,術後改為正常飲食 之組別其 DNA 複製及合成能力並無影響 (Wands et al., 1979)。此現象是由於肝 臟有一自我保護機制,為了預防細胞持續損傷,肝臟會使細胞停止週期運作,並 開始修復 DNA,使細胞於複製前將受損 DNA 還原 (Koteish et al., 2002),不過 在這同一時間代謝產物乙醛也會同時干擾肝細胞週期之 G2 期到 M 期,並開 始進行損害作用 (Clemens et al., 2002; Higgins and Borenfreund, 1986)。
(一) ALD 之相關發炎因子與促發炎激素
細胞激素與發炎、免疫反應及細胞凋亡有關,在外來物的刺激下,許多細胞 例如肝細胞、內皮細胞及星狀膠細胞等皆會分泌細胞激素,並影響周遭細胞及活 化巨噬細胞、單核球細胞和淋巴細胞等參與發炎反應 (Martinez et al., 1992)。酒 精在早期就已有研究明確指出會造成肝臟疾病 (肝炎與肝硬化) (Lelbach, 1976) 和胃潰瘍 (Kvietys et al., 1990),而組織損傷的主因為巨噬細胞分泌之細胞激素如 interleukins-1 (IL-1)、interleukins-6 (IL-6) 和腫瘤壞死因子 (tumor necrosis factor,
12
TNF) 等,這些細胞激素不僅啟動免疫反應,也會參與發炎反應的過程 (Martinez et al., 1992)。
1. 腫瘤壞死因子-α (tumor necrosis factor-α, TNF-α)
TNF-α 參與全身各部位之發炎反應,並為導致急性發炎的主要因子之一,
研究指出許多細胞裡皆有 TNF-α 作用,而肝臟主要是存在於庫氏細胞 (Kupffer cells) 中 (McClain et al., 2004) 。 TNF-α 活 化 後 會 誘 導 粒 線 體 通 透 性 轉 變 (mitochondrial permeability transition, MPT),導致基質脹大、外膜破裂,最後細 胞凋亡並釋放出更多促凋亡因子 (Bernardi et al., 2001)。此外,TNF-α 也會活化 肝細胞中許多壓力活化蛋白激酶 (stress activated protein kinase, SAPK),啟動 p38 mitogen activated protein kinase (MAPK) 及 Jun N-terminal kinase (JNK) (Wajant et al., 2001)。
連續以管餵方式給予大鼠酒精試驗誘發 ALD 的研究發現,肝臟中庫氏細胞 之 TNF-α 含量 隨著 肝損傷程 度而提 高 (Lin et al., 1998) ,Adachi 等 人用 gadolinium chloride 使大鼠庫氏細胞凋亡並同時餵食酒精,發現 TNF-α 及其他 細胞激素含量並無提升,防止肝損傷發生 (Adachi et al., 1994),此外,也可抑制 酒精所造成之脂肪變性 (Jarvelainen et al., 2000)。另外有研究顯示 TNF-α 基因 剔除小鼠餵食大量酒精並不會引起肝損傷 (Kawaratani et al., 2008),因此目前 TNF-α 已被認為是影響 ALD 發展的關鍵細胞激素 (Kitazawa et al., 2003)。
2. 介白素-1β (interleukins-1β, IL-1β)、介白素-6 (interleukins-6, IL-6)
發炎反應是造成 ALD 的原因之一 (McClain et al., 2005),IL-1β 藉由刺激免 疫細胞生長及分化造成發炎反應 (Petrasek et al., 2012),文獻指出 IL-1β 會增加 肝細胞中發炎因子及促脂肪變性趨化因子 monocyte chemoattractant protein-1 (MCP-1),增強巨噬細胞之發炎反應並引起脂肪變性 (Mandrekar et al., 2011;
Miura et al., 2010),而抑制 type I IL-1 receptor (IL-1R1) 則能有效治療 ALD (Petrasek et al., 2012)。IL-6 則屬廣效性細胞激素,負責將急性發炎反應轉為持續 發炎,造成組織損傷 (McClain et al., 1997)。長期飲用酒精之患者及動物試驗皆 可發現血清中 IL-6 含量明顯增加 (Khoruts et al., 1991; McClain et al., 1999)。
3. 誘發型一氧化氮合成酶 (inducible NOS, iNOS)
13
慢性酒精攝取會增加肝臟中一氧化氮 (Nitric oxide, NO) 產生 (Wang et al., 1995),NO 與 superoxide 反應之產物 peroxynitrite 已有研究證實為造成酒精性 肝損傷重要因素之一 (Arteel et al., 1999)。NO 由酵素 NOS 代謝 L-arginine 而 來,細胞中 NOS 又分為三種,分別為 neuronal NOS (nNOS)、inducible NOS (iNOS) 及 endothelial NOS (eNOS) (Stuehr and Griffith, 1992),其中 iNOS 與 ALD 之關係較為密切,在慢性酒精影響下其表現量明顯提高 (McKim et al., 2003),而酒精造成肝炎之研究也指出,此情況下 iNOS 會大量表現,且與肝臟 發炎程度有相同趨勢 (Deng and Deitrich, 2007; Tang et al., 2009)。iNOS 在正常情 況下不表現,但若當體內發生發炎反應時,其表現量受 NF-κB 調控,NF-κB 又 為上游 MAPK 之轉錄產物,而發炎反應與發炎細胞激素相關,因此可得知細胞 激素與受器結合後促使 MAPK 路徑啟動,提升 NF-κB 活性,進一步增加 iNOS 表現量 (Ruimi et al., 2010)。
4. 促分裂素原活化蛋白激酶 (mitogen-activated protein kinases, MAPK)
多種細胞皆有 MAPK 傳遞路徑,活化的 MAPK 從細胞質進入核中,並刺 激許多轉錄因子產生活性,而 MAPK 一般分為三種:胞外訊息調節激酶 (extra -cellular regulated kinase 1/2, ERK1/2)、c-Jun N 端蛋白質激酶 (c-Jun N-terminal kinase, JNK) 及 p38 (Raingeaud et al., 1995)。哺乳動物的 ERK1/2 透過控制生長 因子參與細胞生殖與分化,而 JNK 及 p38 則是有刺激產生時才會被啟動,例 如滲透壓休克、缺血性損傷及發炎細胞激素 (TNF-α、IL-1β) 等 (Kyriakis and Avruch, 2001)。
酒 精 在 腸 道 中 會 改 變 腸 道 通 透 性 , 使 腸 道 內 之 細 菌 內 毒 性 脂 多 醣 (lipopolysaccaride, LPS) 進 入 肝 臟 , 經 巨 噬 細 胞 TLR4 辨 識 後 , 活 化 下 游 ERK1/2 與 p38 傳遞路徑,產生 NF-κB 及促發炎基因表現,造成肝臟發炎 (Mandrekar and Szabo, 2009; Seth et al., 2010)。此外,酒精也會影響巨噬細胞之 NADPH oxidase,使其活性提升進而產生大量 ROS,增加 ERK1/2 及 p38 磷酸 化,提高 TNF-α 表現 (Thakur et al., 2006),另有研究指出 CYP2E1、LPS/TNFα 及 JNK/p38 MAPK 三者具協同作用,導致酒精性肝損傷 (Cederbaum et al., 2015)。幾丁聚醣 chitooligosaccharides (COS) 則是可抑制酒精對 p38、JNK 與 ERK 磷酸化,並提升 Nrf-2 來達到保肝作用 (Luo et al., 2014)。
14 5. NF-κB
NF-κB 是位於核內的轉錄因子,調節各種基因表現,例如 iNOS、COX-2 等 細胞激素,而當體內出現細胞凋亡、腫瘤形成、自體免疫疾病及發炎反應的情況 時,皆會發現 NF-κB 表現 (Lawrence et al., 2001)。NF-κB 為同源二聚體 p50 及 異源二聚體 p65 之複合物 (Baldwin, 1996),正常情形下 NF-κB 與抑制蛋白 Iκ -Bα 結合,形成不活化複合物存在於細胞質中,使 NF-κB 無法進入核中調控相 關基因 (Miyamoto and Verma, 1995),而當有發炎情形時,IKK-α complex 便會 活化,並進一步降解 IκBα 形成 IκB,釋放 p65/p50 二聚體至核內啟動轉錄 NF-κB 機制 (Baldwin, 1996; Zandi et al., 1997)。
餵食雄性 Wistar 大鼠酒精液態飼料誘發肝損傷試驗結果發現,活化 NF-κB 會使促發炎激素 mRNA 表現量提高 (Jokelainen et al., 2001);探討酒精性肝損傷 中 iNOS、eNOS、NF-κB 及 TNF-α 相互關係之研究表明,酒精會使雌性 SD 大 鼠肝臟發炎,並增加 iNOS、NF-κB、TNF-α 表現,而 eNOS 活性雖降低但其 mRNA 表現量不受影響 (Yuan et al., 2006)。薑黃素有效預防雌性 SD 大鼠經四 周餵食酒精誘發肝損傷之原因為其能顯著降低氧化壓力與抑制 NF-κB 活性,改 善初期肝損傷 (Nanji et al., 2003)。
6. 第二型環氧化酶 (cyclooxygenase-2, COX-2)
COX 負 責 催 化 花 生 四 烯 酸 (arachidonic acid) 轉 變 為 前 列 腺 素 H2 (prostaglandin H2) 與調控多種生物活性前驅物如前列腺素 E2 (prostaglandin E2)、
前列環素 (prostacyclin) 和血栓素 A2 (Thromboxane A2) 等 (Hawkey, 1999;
Picot et al., 1994)。COX 分為兩種型態:COX-1 在哺乳細胞中作為維持生理狀 態之前列腺素合成;COX-2 正常狀態時為低度表現或不表現,經發炎細胞激素 刺激後才活化 (Mitchell et al., 1995; Smith et al., 1996; Vane et al., 1994)。
COX-2 經巨噬細胞及內皮細胞之發炎因子誘導而成,過度發炎所造成之疾 病如類風濕關節炎、慢性肝炎或是肺纖維化等,皆可發現其大量表現 (Coker and Laurent, 1998; Isomaki and Punnonen, 1997; Tilg et al., 1992),此外,以酒精管餵大 鼠研究中也發現酒精性肝損傷之形成為酒精刺激肝臟 Kupffer cell 大量表現 COX-2,造成脂質過氧化及壞死性炎症 (Nanji et al., 1997)。
(二) ALD 之抗發炎因子
15
1. 過氧化體增生劑活化受體-γ (peroxisome proliferator activated receptor-γ, PPAR-γ)
PPAR-γ 為 PPARs 三種異構體之一,另外兩個異構體分別為 PPAR-α 及 PPAR-β (Evans, 1988),PPAR-γ 主要調節細胞分化、凋亡、脂質代謝及發炎反應 (Houseknecht et al., 2002),而研究指出當體內發生發炎反應時,單核細胞分化為 巨噬細胞過程會誘導 PPAR-γ 表現 (Chinetti et al., 2000; Tontonoz et al., 1998)。
許多文獻顯示,當 PPAR-γ 配體如 15d-PGJ2、TZDs 及 NSAIDs 大量表現時,
PPAR-γ 會抑制 iNOS、基質金屬蛋白酶-9 (matrix metalloproteinase-9, MMP-9) 與 scavenger receptor A 轉錄 (Ricote et al., 1998),此外,體外試驗也證實 PPAR-γ 亦可抑制細胞激素 (TNF-α、IL-1β 和 IL6) (Chinetti et al., 2000; Debril et al., 2001;
Gelman et al., 1999),可能具良好抗發炎之能力 (Castrillo and Tontonoz, 2004;
Daynes and Jones, 2002)。Zhao 等人發現,當大鼠罹患酒精性肝纖維化的同時,
PPAR-γ 表現量大幅降低,推測 PPAR-γ 在酒精所引起之肝臟發炎、壞死及纖維 化中有著重要作用 (Zhao et al., 2004),而以酒精及高油脂飲食誘導大鼠脂肪肝研 究也發現肝臟發炎會導致 PPAR-γ 表現量下降,因此認為透過配體激活 PPAR-γ 可做為治療脂肪肝之方法 (Zhao et al., 2004)。
三、ALD 之治療方式
目前治療 ALD 除了改變生活型態之外,也可藉由藥物例如 naltrexone 和 acamprosate 協助戒酒 (Bouza et al., 2004),或是額外補充鋅及蛋白質營養素可改 善或減輕 ALD 之情形 (Kang and Zhou, 2005; McCullough and O'Connor, 1998)。
而 於 臨 床 上所 使 用 之藥 物 有 糖 皮質 激 素 (glucocorticosteroids) 、己酮可可鹼 (pentoxifylline)、抗氧化劑與直接抗纖維化物質等,其方法大多為直接改善肝臟 損傷和促進肝臟修復 (Frazier et al., 2011; Lucey, 2009; Raetsch et al., 2002)。氧化 壓力被認為是在 ALD 中扮演重要的角色,因為酒精造成之脂質過氧化、ROS 產 生和內源性抗氧化能力衰竭皆為氧化壓力所導致 (Dey and Cederbaum, 2006)。
Silymarin 為乳薊草萃取物中多種黃酮素的總稱,具良好的抗氧作用,也是治療 肝臟疾病最常使用藥物之一。文獻證實 silymarin 具清除自由基與增加肝臟抗氧 化酵素之能力,防止脂質過氧化,並同時促進肝細胞修護與再生 (Soto et al., 2003)。在許多動物模式與臨床研究中 silymarin 均有保護肝臟及改善肝損傷之 功能,而在目前肝臟疾病之應用包括抗發炎、抗氧化、抗纖維化、免疫調節以及
16
抑制酒精誘導之肝毒性 (Del Prete et al., 2012; Lieber et al., 2003; Song et al., 2006)。
四、酒精性脂肪肝 (alcoholic fatty liver, AFL)
正常肝臟中之脂質約少於 5%,若肝臟脂質堆積介於 5-33% 定義為輕度脂 肪肝,33-66% 屬中度脂肪肝,重度則是大於 66%,且肝臟中主要囤積之脂肪為 三酸甘油酯 (triglycerides, TG) (Brunt et al., 1999),其生成代謝路徑如圖 1-4 所 示。組織病理學上罹患酒精性脂肪肝之切片中會有約 82% 出現中度至重度脂肪 浸潤、76% 具有 Mallory 小體、68% 會產生硬化性透明壞死 (sclerosing hyaline necrosis),另外還會有約 97% 之切片會出現氣泡狀的肝細胞變性合併多型核細 胞浸潤之情形 (MacSween and Burt, 1986)。
酒精經代謝會產生大量之 NADH,而 NADH 會增加甘油磷酸鹽去氫酶 (glycerophosphate dehydrogenase, GPDH) 活性,進一步提升 glycerol-3-phosphate (glycerol 3-P),glycerol 3-P 提供了脂質合成所需之 glycerol,因此含量提高會促 進肝臟脂質合成 (Andersen and Holub, 1976),此外,Iseri 等人研究也指出酒精 會破壞氧化脂肪酸的粒線體結構,使氧化能力大幅下降,代謝酒精過程造成之氧 化還原狀態不平衡狀態會使氫離子大量增加,降低脂肪酸氧化作用,而來不及被 氧化之脂肪酸便會進入血液中回流至肝臟堆積 (Iseri et al., 1966)。
正常情況下累積在肝臟裡的 TG、膽固醇與磷脂質會被 lysosomes 中的脂解 酵素進行水解作用,進而被身體利用,但長期攝取酒精除了會使脂解酵素失活外 (Fowler and De Duve, 1969),水解作用也會被影響 (Takeuchi et al., 1974),且也有 研究顯示酒精會使膽汁分泌量減少,所以在肝臟中轉變為膽酸的膽固醇也會隨之 減少,而未被利用的 TG、膽固醇與磷脂質便會累積在肝臟,間接形成脂肪肝 (Maddrey and Boyer, 1973)。
酒 精 性脂 肪肝 之分 子 機轉 如圖 1-5,根 據 研究 顯示 酒精 會抑 制 AMPK (AMP-activated protein kinase),影響脂肪合成及降解之間平衡的關鍵代謝產物,
例如 malonyl coenzyme A 和長鏈 acyl-coenzyme A (You et al., 2004)。酒精也會誘 導脂質合成酵素 (lipogenic enzymes) 過量表現,進而抑制 PPAR-α (Sozio and Crabb, 2008)、提升固醇類轉錄因子 SREBP (sterol regulatory element-binding protein) (Galli et al., 2001),最終造成酒精性脂肪肝。
17 圖 1-4 肝臟中三酸甘油酯代謝合成路徑
Fig. 1-4 Metabolic pathways leading to the synthesis of TG in liver (Postic and Girard, 2008)
18
圖 1-5 酒精造成脂肪肝及脂毒性之主要途徑
Fig. 1-5 Schematic diagram of the major pathways of alcoholic fatty liver and lipotoxicity (Purohit et al., 2009)
19 (一) 肝臟脂質代謝與合成相關蛋白
1. 腺苷酸活化蛋白激酶 (AMP-activating protein kinase, AMPK)
AMPK 是由三種次單位形成的三聚體,分別為 α、β 及 γ (Hardie and Carling, 1997)。哺乳動物有許多部位皆有 AMPK,例如骨骼肌、肝臟、脂肪細 胞等 (Misra and Chakrabarti, 2007),其活化路徑有兩種,一為 AMP/ATP 比值升 高時,如長時間運動 (Hutber et al., 1997)、骨骼肌電位差 (Winder and Hardie, 1996)、心肌局部缺血 (Kudo et al., 1995)、熱休克 (Corton et al., 1994) 等,二是 當肌酸/磷酸肌酸 (creatine/ phosphocreatine, pCr) 比值增加也會刺激 AMPK 產 生 (Hardie and Carling, 1997)。
AMPK 於細胞內主控調節代謝、氧化壓力和能源支配,也能增加醣解與脂 肪酸氧化作用,以及調控脂肪酸、膽固醇和蛋白質的合成 (Sozio and Crabb, 2008)。
被刺激之 AMPK 與 AMP 結合後會提升細胞能量,抑制脂肪及膽固醇合成等代 謝機制。反之,若 AMP/ATP 比值降低,AMPK 便不具活性,而細胞則繼續進 行脂肪及膽固醇的合成,由此可知 AMPK 對於脂質代謝與合成是相當重要的 (Misra and Chakrabarti, 2007)。而調控 AMPK 之關鍵因子為丙二醯輔酶 A ( malonyl CoA) 與粒線體中之長鏈醯基輔酶 A (acyl-coenzyme A),其透過調控固 醇類調節結合蛋白-1C (SREBP-1C) 以及抑制乙醯輔酶 A 羧化酶 (acetyl-CoA carboxylase, ACC)和 3-羥基-3-甲基還原酶 (3-hydroxy-3-methylglutaryl, HMG- CoA reductase) 來直接影響脂質合成 (Sozio and Crabb, 2008)。
酒精與肝臟中 AMPK 之相互關係已有許多研究證實,Javier Garcı´a 等人發 現酒精會使 AMPK 活性降低,造成 AMPK 磷酸化受阻並增加 ACC 活性,大 幅提升 malonyl-CoA 含量,影響脂肪酸代謝並有利於 TG 累積在肝臟形成酒精 性脂肪肝 (AFL) (Garcia-Villafranca et al., 2008)。亦有研究指出活化 AMPK 可促 進脂肪酸氧化並降低脂肪形成,但攝入酒精會抑制其活性,進而助於酒精性脂肪 變性 (Molina, 2008)。在脂質生合成代謝路經中,AMPK 透過控制 SREBP-1、
ACC 與過氧化體增殖活化受體-α (peroxisome proliferator activated receptors, PPAR-α) 達到預防脂肪肝的形成 (You and Crabb, 2004)。
2. 固醇類調節結合蛋白 (sterol regulatory element-binding proteins, SREBPs) SREBPs 是調控脂肪酸、三酸甘油酯與膽固醇合成的轉錄因子,未活化時存 在於內質網及核模上,一旦被活化就會透過 SREBP cleavage-activating protein
20
(SCAP) 從核膜上運輸至高基氏體中,並與特定蛋白作用後進入細胞核內進行固 醇類調節與合成 (Sozio and Crabb, 2008)。目前已知有 SREBP-1a、SREBP-1c 與 SREBP-2 三種 SREBPs (Shimano, 2001)。
SREBPs 為 AMPK 之下游蛋白,而 SREBP-1 負責 TG 及脂肪酸合成 (Shimomura et al., 1999),研究顯示當小鼠肝臟過量表現 SREBP-1 時,其脂肪酸 合成速率足足提高四倍,並影響下游脂質生成基因例如 fatty acid synthase (FAS) 和 glycerol-3-phosphate acyltransferase (GPAT) 等表現 (Shimomura et al., 1998)。
酒精會影響肝臟中 PPAR-α、SREBP-1 及 AMPK,抑制脂肪酸氧化與促進 TG 合成,並同時產生極低密度脂蛋白 (very low density lipoproteins, VLDL),最終導 致酒精性肝損傷 (Rasineni and Casey, 2012)。SREBP-2 則是負責調控膽固醇與低 密度脂蛋白受體 (low density lipoproteins receptor, LDLR) (Horton et al., 2002)。
Wang 等人用酒精誘導大鼠脂肪肝及肝損傷之研究發現酒精會不斷促使肝臟中 SREBP-2 活化及增加 3-hydroxy-3-methyl-glutaryl-CoA (HMG-CoA) 表現量,提 升膽固醇合成速率增加膽固醇含量,此外,也會降低 LDLR,形成高血脂症 (Wang et al., 2010)。
3. 過氧化體增殖劑活化受體-α (peroxisome proliferator-activated receptors-α, PPAR-α)
PPAR-α 存在於棕色脂肪、肝、腎、胃和心臟等組織中,屬於 peroxisome proliferator-activated receptors (PPARs) 家 族 之 一 , 而 PPARs 家 族 另 外 還 有 PPAR-β (PPAR-δ) 與 PPAR-γ ,PPAR-β 於腎及肺表現量最高,PPAR-γ 則是主 要表現於脂肪組織及免疫系統中 (Braissant et al., 1996; Desvergne and Wahli, 1999)。
PPAR-α 於肝臟中負責脂肪酸運輸及氧化,活化後之 PPAR-α 與 retinoid X receptor (RXR) 結合會提升脂肪酸 β-oxidation 作用,並調節長鏈 acyl-CoA synthetase 基因 ,進而降低 TG 含量,同時增加細胞抗氧化壓力酵素,減少細 胞凋亡 (Motojima et al., 1998; Sozio and Crabb, 2008)。此外,PPAR-α 也透過調 控脂肪酸氧化相關基因,例如控制脂肪酸運輸之 carnitine palmitoyl transferase-I -1 (CPT-1) 及過氧化小體中脂肪酸 β-氧化作用 (β-oxidation) 之速率限制步驟的 關鍵酵素 ACOX (Acyl-CoA oxidase) 等 (Kohjima et al., 2007)。文獻指出,給予 高油脂飲食誘發粥狀動脈硬化小鼠 PPAR-α 活化劑可有減緩主動脈弓 (aortic
21
arch) 50%、胸腔主動脈 (descending thoracic aorta, DTA) 及腹主動脈 (abdominal aorta) 90% 之粥狀硬化程度,也可抑制小鼠腹腔巨噬細胞泡沫細胞的形成,降低 50% 高膽固醇血症 (Li et al., 2004)。酒精會使小鼠體內 RXR 降低,進而影響 PPAR-α 活化,使脂肪酸氧化無法作用 (Galli et al., 2001),但若酒精和 PPAR-α 活化劑同時餵食則是可以明顯防止酒精性脂肪肝之情形 (Fischer et al., 2003)。
Microsomal triglyceride transfer protein (MTP) 負責運輸極低密度脂蛋白 (very low density lipoproteins, VLDL) 至肝臟外,而 MTP 由 PPAR-α 所調控,因此,
當酒精抑制 PPAR-α 活化,便也停止 MTP 作用,使 VLDL 輸出受限,並在肝 臟中累積 (Nanji et al., 2004; Sugimoto et al., 2002)。Nakajima 等人以酒精餵食 PPAR-α knockout 小鼠以分析酒精性肝損傷分子機制,發現缺乏 PPAR-α 小鼠與 wild-type 相比其肝細胞損傷與肝腫大情形皆嚴重許多,並減少許多抗氧化酵素 如 GPx、SOD 等,還會提高發炎因子 TGF-α 及 TGF-β1,發展為酒精性肝纖維 化 (Nakajima et al., 2004)。
4. 乙醯輔酶 A 羧化酶 (acetyl-CoA carboxylase, ACC)
AMPK 作用會進一步抑制 ACC 活性,而 ACC 之功能是將 acetyl-CoA 催 化為 malonyl-CoA,其產物會促使脂肪生成 (Wakil et al., 1983) 及 β-oxidation (McGarry and Brown, 1997),因此通常被看作為脂肪酸合成限速酶 (You and Crabb, 2004)。在哺乳動物體內 ACC 具兩種型態,個別存在於不同組織中且生 理作用也大不相同,ACC1 (ACCα) 於細胞質裡,並廣泛分布在肝臟和脂肪組織 中,參與脂肪生成;ACC2 (ACCβ) 位於心肌與骨骼肌之粒線體,肝臟含量較少 (Abu-Elheiga et al., 2000)。不過 Mao 等人研究指出剔除 ACC1 基因小鼠雖然可 以降低肝臟中 malonyl-CoA、脂肪酸合成與 TG 含量,但並無法預防高油脂飲 食所產生之肥胖與脂肪肝 (Mao et al., 2006)。而後,Harada 等人發現肝臟缺少 ACC1 小鼠其 malonyl-CoA 含量並未受影響,證實 ACC2 參與了 malonyl-CoA 合成 (Harada et al., 2007)。由上述研究可得知 ACC 分別有兩種調節脂肪酸合成 路徑,ACC1 是促進脂肪酸合成之角色,而 ACC2 所產生之 malonyl-CoA 透過 抑制 carnitine palmitoyl-CoA transferase-I 調控脂肪酸氧化 (Brownsey et al., 2006)。
而在酒精的刺激下會抑制 AMPK 活化,使 ACC 活性增加,促使肝臟進行脂肪 酸合成作用,並將游離脂肪酸代謝成 TG 之形式累積在肝臟中 (Yin et al., 2007)。
22 5. 脂肪酸合成酶 (fatty acid synthase, FAS)
FAS 催化 malonyl-CoA 轉變為棕櫚酸 (palmitate) (Alberts et al., 1975),為合 成 脂 肪 酸 之 最 後 步 驟 , 因 此 被 認 為 是 肝 臟 中 重 要 的 脂 肪 生 成 決 定 因 子 (Abu-Elheiga et al., 2005)。FAS 存在於不同組織中,其功能也大不相同,它會將 肝臟中之醣類轉為脂肪酸,使脂肪生成並儲存於肝臟中,而乳腺細胞中之 FAS 則是可合成嬰兒較容易吸收的脂肪酸 (Wakil et al., 1983)。研究顯示酒精會使 SREBP-1 表現提升,下游因子 FAS 表現量也因此提高,藉此助於 TG 生成並 累積於肝臟中,導致酒精性脂肪肝 (You et al., 2002)。
第三節 紅麴菌
紅麴又名赤麴或丹麴,日本稱為 beni koji 或 anka koji,歐洲則以中國紅米 (red Chinese rice) 稱之 (陳,2003),在亞洲地區,紅麴於飲食、醫藥與工業上之 應用已有千年的歷史,西元 1596 年,明代李時珍所撰之《本草綱目》中對紅麴
功效記述如下:「主治消食活血,健脾燥胃」。明末清初,鄭成功光復台灣後,廣
招漢人來台發展開墾,而紅麴的出現,相傳便是在這時期,自大陸浙江、江蘇、
江西與福建一帶渡海來台的製酒匠人所傳入,而後到了日本統治時期,台人於樹 林設立樹林造酒公司,並開始大量生產「金雞」和「黃雞」兩種紅酒,也就是後 來知名的紅露酒。時至今日,隨著微生物的應用技術愈趨完熟與國人對養生概念 的日趨重視,因此,在國內、外,紅麴於保健食品上的應用皆有不錯的發展,也 在在證明紅麴乃是一個藥食同源的最佳代表。
一、紅麴菌之菌種特性
紅麴菌之生活史如圖 1-6 所示,紅麴菌為雌雄同體 (homothallic),其營養 菌絲為無色、褐色或紅色,且呈不規則狀分散,內含大型液泡、微小體與粒線體,
具有橫隔,可由菌絲末端形成一個大型的有性厚壁子囊果形有性生殖,或散佈分 生胞子 (conidium) 行無性生殖。當行有性生殖時,位於菌絲頂端之精子器 (antherdium) 會延長為一多核管狀細胞,此時,雌性母細胞將分裂為受精毛 (trichogyne) 與造囊果 (ascogonium),而精子器之核透過受精毛進入造囊果中並
23 圖 1-6 紅麴菌之生活史
Fig. 1-6 Life cycle of Monascus species. (蘇等,1970) 1, 2:子囊孢子形成營養菌絲
3→7:生殖器官之形成與造囊菌絲之發育 8, 9:成熟之子囊果
10:單一細胞分生孢子 (one cell conidia) 之無性生殖
24
結合,而後形成一直徑約 20-40 μm 的子囊果,待成熟後,子囊胞子將自子囊果 之裂口處釋出,並繼續其生活史 (蘇等,1970)。
二、紅麴菌之分類
紅麴菌可於自然界中之穀類、澱粉、新鮮牧草、泥土、河川表面沉澱物與松 樹根組織中發現,紅麴菌是 1884 年由法國學者 Van Tieghem 自汙染穀類及農 產品等作物中分離而得菌種,繼而開啟紅麴菌之命名與分類 (陳等,2003)。紅 麴 菌 在 分 類 學 上 屬 於 真 菌 界 (Fungi) 、 子 囊 菌 門 (Ascomycota) 、 子 囊 菌 綱 (Ascomycetes) 、 散 囊 菌 目 (Eurotiales) 、 紅 麴 菌 科 (Monascaceae) 、 紅 麴 菌 屬 (Monascus)。紅麴菌特徵為具有橫隔菌絲 (septate hypha),且末端可產生有性的 厚壁子囊,而分類學家依此特徵遂將其歸類為子囊菌門 (林,1983;林,1995)。
已知目前文獻上被命名的紅麴菌約有 20 個種,起初紅麴菌之分類是依據種源、
培養基型態與發酵特性等特徵來命名;1936 年佐藤喜吉則根據菌絲有無著色粒 子與菌叢顏色,進而將紅麴菌分為白色與有色兩大類以及五個亞類;1983 年,
Hawskworth 與 Pitt 兩人以紅麴菌種於洋菜固態培養基之生長速度、菌叢顏色、
閉囊果與分生胞子大小、顏色等特性將紅麴菌重新歸類並分為三種,分別是 Mruber、M. purpureus 與 M. pilosus (Hawksworth and Pitt, 1983)。而後到了 1995 年,紅麴在分類上又另分出兩種菌種,分別為 M. pallens 及 M. sanguineus (Cannon et al., 1995)
三、紅麴於預防醫學之應用 (一) 血脂調節之相關研究
在東南亞,紅麴菌應用於食品發酵已有好幾世紀,近年來,紅麴發酵產品已 逐漸發展為預防心血管疾病之保健食品 (Lee and Pan, 2012b)。而紅麴降血脂之.
功效早在西元 1999 年就已受到證實,研究人員於人體雙盲試驗中探討紅麴米對 於血脂之影響,研究結果顯示,在為期 12 周的試驗日程中,服用紅麴之患者與 對照組相比,前者血中 TC 與 LDL-C 含量於試驗第 8 周,已顯著低於對照組,
且服用紅麴米之患者,其各項肝功能指標與對照組均無明顯差別,也開啟日後紅 麴對於調控血脂之相關研究 (Heber et al., 1999)。2000 年 Wang 等人以高果糖 飲食誘導大鼠產生高 TG 血症,實驗期間輔以紅麴米的餵食,結果顯示紅麴米 能顯著降低高 TG 血症大鼠血清中 TC、TG、LDL-C 與 VLDL-C 含量,並提
25
高肝臟中脂解酵素活性 (Wang et al., 2000)。2003 年 Wei 等人探討紅麴米對高膽 固醇血兔降血脂與抗動脈粥狀硬化效果之影響,研究結果顯示紅麴米雖無法顯著 降低血清中 TC、TG 與 LDL-C 含量,但卻可提升血清中 apo A 與降低 apo B 含量,並改善肝臟中脂質過氧化程度 (Wei et al., 2003)。2006 年 Lee 等人將 M.
purpureus HM105 紅麴菌株進行突變並篩選出 M. purpureus NTU568,接著以 M.
purpureus NTU568 餵食經高膽固醇飲食誘發高血脂症之倉鼠,從結果可知突變
後的新菌株 M. purpureus NTU568 對血清及肝臟中 TC 與 TG 含量均有顯著 降低效果,亦能降低血清中 LDL-C 含量,重要的是,餵食經 M. purpureus NTU568 發酵之紅麴米還可提升血清中 HDL-C 含量,且對肝臟也不具毒性 (Lee et al., 2006a)。2007 年,Lee 等人發現以山藥作為紅麴固態發酵基質比傳統 紅麴米具有更良好降血脂預抗動脈粥狀硬化之功效,研究結果表明,餵食 0.5 倍 之紅麴山藥便可達到餵食 1 倍紅麴米所提供的降 TC 與 TG 效果,而餵食 1 倍之紅麴山藥所降低的 TC 含量更顯著高於餵食 1 倍紅麴米,另外,1 倍紅麴 山藥相比於 1 倍紅麴米,前者對於血液中抗氧化酵素 SOD 與過氧化氫酵素活 性有更佳的提升效果,最後在引起動脈粥狀硬化的脂質斑塊面積方面,1 倍紅麴 山藥抗動脈粥狀硬化之能力更顯著高於 1 倍之紅麴米 (Lee et al., 2007a)。此外,
紅麴發酵之大豆萃取物不僅可降低高血脂症大鼠血清中 TC、TG 與 LDL-C 含 量,還可抑制 3-羥基-3-甲基戊二酸醯輔酶還原酵素 (3-hydroxy-3-methylglutaryl coenzyme A reductase, HMG-CoA reductase) 活性 (Pyo and Seong, 2009)。本實驗 室之過去研究發現,以 DOW 所發酵的紅麴山藥對於降低血清及肝臟中 TC 含 量的能力不僅高於 ROW 所發酵的紅麴山藥。以 DOW 作為發酵水分來源的紅 麴山藥更具有提升血清中 HDL-C 含量的效果,另外,DOW 所發酵之紅麴山藥 與 ROW 發酵之紅麴山藥相比,前者在減緩脂質過氧化程度與降低心臟主動脈 脂質斑塊堆積方面具有更顯著之效果 (Lee et al., 2011b)。最近研究指出,富含輔 酶 Q9 與 Q10 之紅麴發酵混合榖物萃取物可明顯降低血清中 TC、TG 與 LDL-C 含量,對於脂質過氧化程度也具降低作用,此外,攝取紅麴發酵混合榖 物萃取物還可提升高血脂症大鼠肝臟、心臟與血漿中之輔酶 Q9 與 Q10 含量 (Pyo and Seong, 2013),這或可解決因長期服用含高量 monacolin K 之食品而引 起如橫紋肌溶解症等副作用的問題。以上研究結果在在肯定紅麴具有良好的血脂 調節能力。